
Hybrids, the results of successful breeding between two parents of different species, occur frequently in nature, yet are perhaps most familiar to us when they result from human intervention. We encounter in our intentionally cultivated hybrids the utility of the mule, the stateliness of the London plane tree, and the sensuous smells and tastes of myriad vegetables and fruits, including broccolini and the tangelo. These remarkable examples are of our own making, but hybridization between closely related species is perhaps the norm rather than the exception in nature. Though hybrid offspring are sometimes sterile and can be visually distinct from their parents (like mules), they are just as likely to be fertile and to pass unnoticed by us. These cryptic hybrids, diagnosable only through genetic testing, breed with each other or with individuals of their parent species (in a process called backcrossing), giving rise to new hybrid progeny. Over generations, such interbreeding consolidates novel hybrid traits, sometimes leading to the formation of new species. Because what counts as a species is, after all, merely conventional, it could be said that we humans, the descendants of interbreeding between Neanderthals and early Homo sapiens, are just as much hybrids as the most luscious of tangelos. Perhaps foremost among the natural world’s “hopeful monsters”—a term that geneticist Richard Goldschmidt coined in 1940 for evolutionary transformations that occur through rare but large-scale mutation—hybrids often captivate and delight human observers.1
Yet hybridization does not always precipitate the formation of a new species. In natural populations, hybrids are frequently formed, only to be subsumed, through backcrossing, into their parental stock. This process—called introgression—results in the enrichment of the gene pool of the predominant species with genetic material from close relatives. So, in the case of oaks—described as particularly problematic for the biological species concept due to their wanton tendency to hybridize (Burger, 1975)— we might say that there is evidence of red oak (Quercus rubra) introgression into a stand of northern pin oaks (Q. ellipsoidalis). These pin oaks will still be pin oaks, but perhaps with some hidden genetic diversity and leaves or bark that look, well, just a little bit different. The ubiquity of such situations has led biologists to formulate the idea of hybrid complexes or zones: sets of species or populations in which rampant interbreeding has produced a messy gradient of similar organisms, rather than discrete sets. Our cultivated citruses represent one such complex, in which ten progenitor species in southeast Asia and Australia have given rise, through hybridization, to dozens of domesticated taxa (Wu et al., 2018).


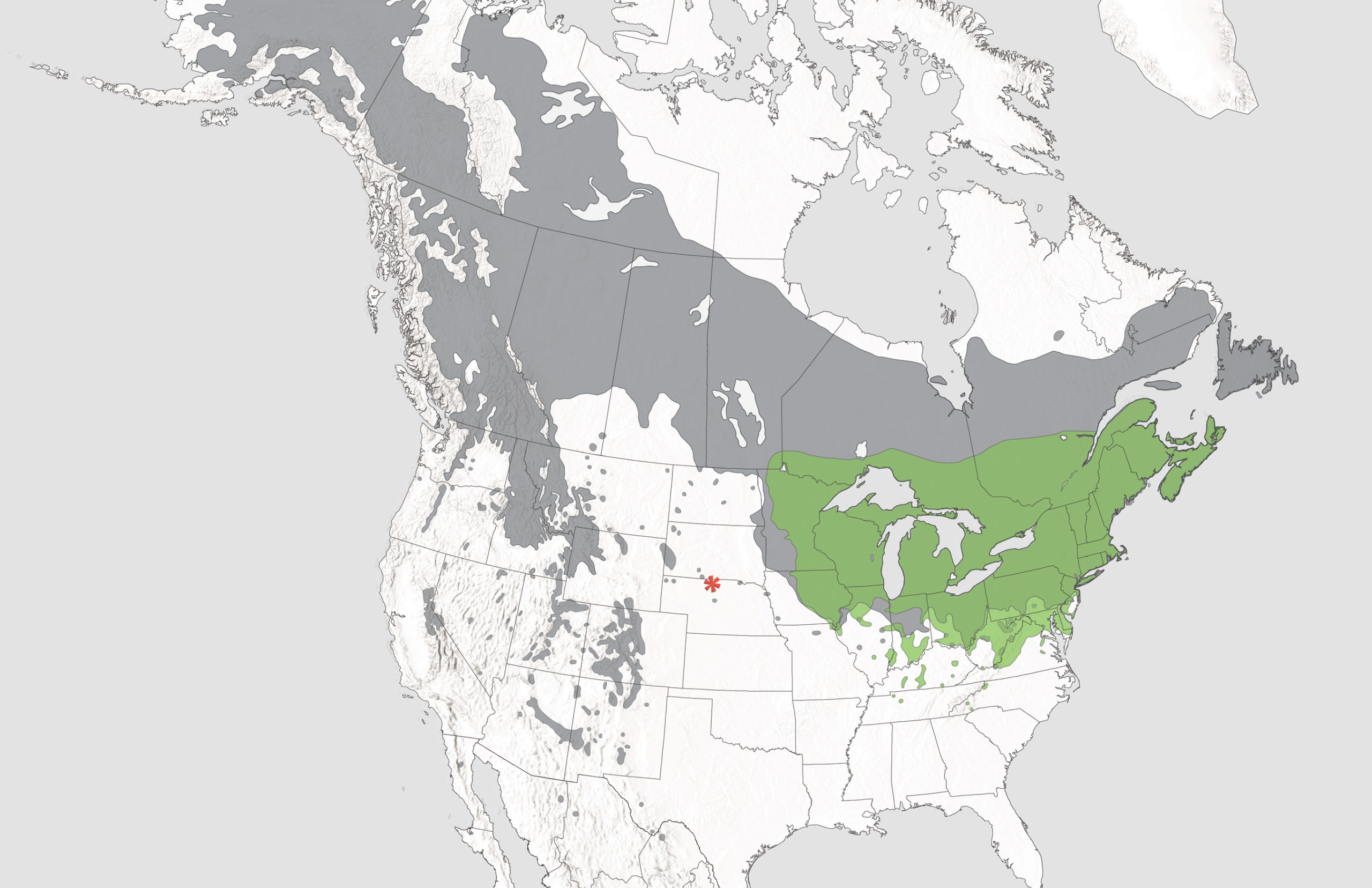
And so, when I began my own foray into the world of hybrid aspens (Populus), I risked wading into a thicket of biological questions that could have been difficult or impossible to resolve. Fortunately, I was a first-year graduate student, a neophyte far more optimistic than I am now when it comes to tackling a new project. What follows is a story of a journey through which a team of ecologists and evolutionary biologists, myself included, tried to track down the truth about a putative hybrid. To do so, we traveled throughout the Midwest and dug deep into the natural history of the Niobrara River Valley, a relictual ecoregion left behind by the retreat of glaciers at the end of the last Ice Age.
A biotic crossroads: The Niobrara River Valley
Heading into the Nebraska Sandhill region along the state’s border with South Dakota, new visitors might be surprised to plunge from cornfields and pastures stretching as far as the eye can see into forested canyons hugging a cool, inviting river. Originating on the eastern edge of Wyoming, the Niobrara River runs from west to east across the northern quarter of Nebraska before being subsumed in the Missouri River in the northeastern corner of the state. The canyons found between the river’s banks and the surrounding matrix of arid short- and mixed-grass prairie constitute the Niobrara River Valley.
For an eager observer, this valley presents more than just a respite from Nebraska’s hot, dry uplands. Rather, as Midwestern botanist Charles Bessey first observed in 1887, the Niobrara River Valley is a “meeting-place for two floras,” a unique location in North America in which East and West comingle. Born and raised in Arizona, I had spent about five years living in the Midwest by the time I first visited the valley. When I arrived by car from St. Paul, Minnesota, where I had just begun my doctoral studies as a plant ecologist, it felt like I was seeing old friends again after a long absence. Most noticeably, stands of ponderosa pine (Pinus ponderosa), a decidedly western species, greet visitors to the Niobrara. These pines are among over a dozen western vascular plant species whose distributions extend all the way into Nebraska, following the biotic east-west highway formed by the Niobrara River. A second look confirmed that this traffic moved in both directions: bur oaks (Quercus macrocarpa) and silver maples (Acer saccharinum) made unusual appearances for species that generally cannot be found in great abundance further west than the meeting of the prairies and forests in Minnesota and Iowa. It also became clear that the cooler, north-facing slopes of the valley, in particular, offered suitable habitat for species generally found further north, including cosmopolitan but drought-intolerant paper birch (Betula papyrifera). The same pattern— a confluence of biota typical of the montane West, the deciduous forests of the East, and the boreal forests of the North—holds for herbaceous plants, insects, and vertebrates as well (Kaul et al., 1988).
Of aspens, poplars, popples, and cottonwoods
Any exhaustive flora of the Niobrara is bound to mention aspens, meaning species of the genus Populus. These trees go by a plethora of common names: aspens, poplars, popples, and cottonwoods—names which do not neatly map on to the current phylogenetic characterization of six sections within the genus (Hamzeh and Dayanandan, 2004). In North America, native species from section Populus, and some from Tacamahaca, are referred to as “aspens,” “poplars,” or “popples.” These include the ubiquitous quaking aspen (P. tremuloides), its close relative bigtooth aspen (P. grandidentata), and cold-tolerant basalm poplar (P. balsamifera). The species we know as “cottonwoods” are restricted to Aigieros and also to Tacamahaca, and these include, most famously, eastern cottonwood (P. deltoides, one of the largest trees east of the Mississippi), Fremont cottonwood out west (P. fremontii), and northwestern black cottonwood (P. trichocarpa), the first tree species to have its genome sequenced. Most species in the genus share traits with each other and with other members of the willow family (Salicaceae): they are dioecious, meaning that male and female flowers are borne on separate trees and have simple leaves and relatively short lifespans (often less than one hundred years). Many species have circumboreal distributions; they are generally cold tolerant, but vulnerable to hot and dry conditions.
Aspens writ large have captured the imagination of botanists and the general public alike by virtue of their propensity for perpetuation through vegetative means. Most importantly, this means that a given aspen tree can, regardless of sex, produce new, genetically identical clones of itself. These new stems, often called “suckers,” emerge from rhizomes, underground stems that spread in parallel to the soil surface. When a rhizome’s buds encounter moist, warm conditions, a small shoot heads upward, sprouting leaves and emerging from the soil as though a seed had germinated in that exact spot. But these suckers grow faster than a typical seedling ever could, drawing on resources from their parental plant and bypassing seedling-hood in a mad dash for growth. These clonal offspring can, over time, become separated from their parents through the decay of the rhizome, but they remain genetic clones, such that what often appears to be a stand of aspen trees is, in reality, a single individual, connected, to varying extents, underneath the soil.
The mystery of the Smith Falls aspens
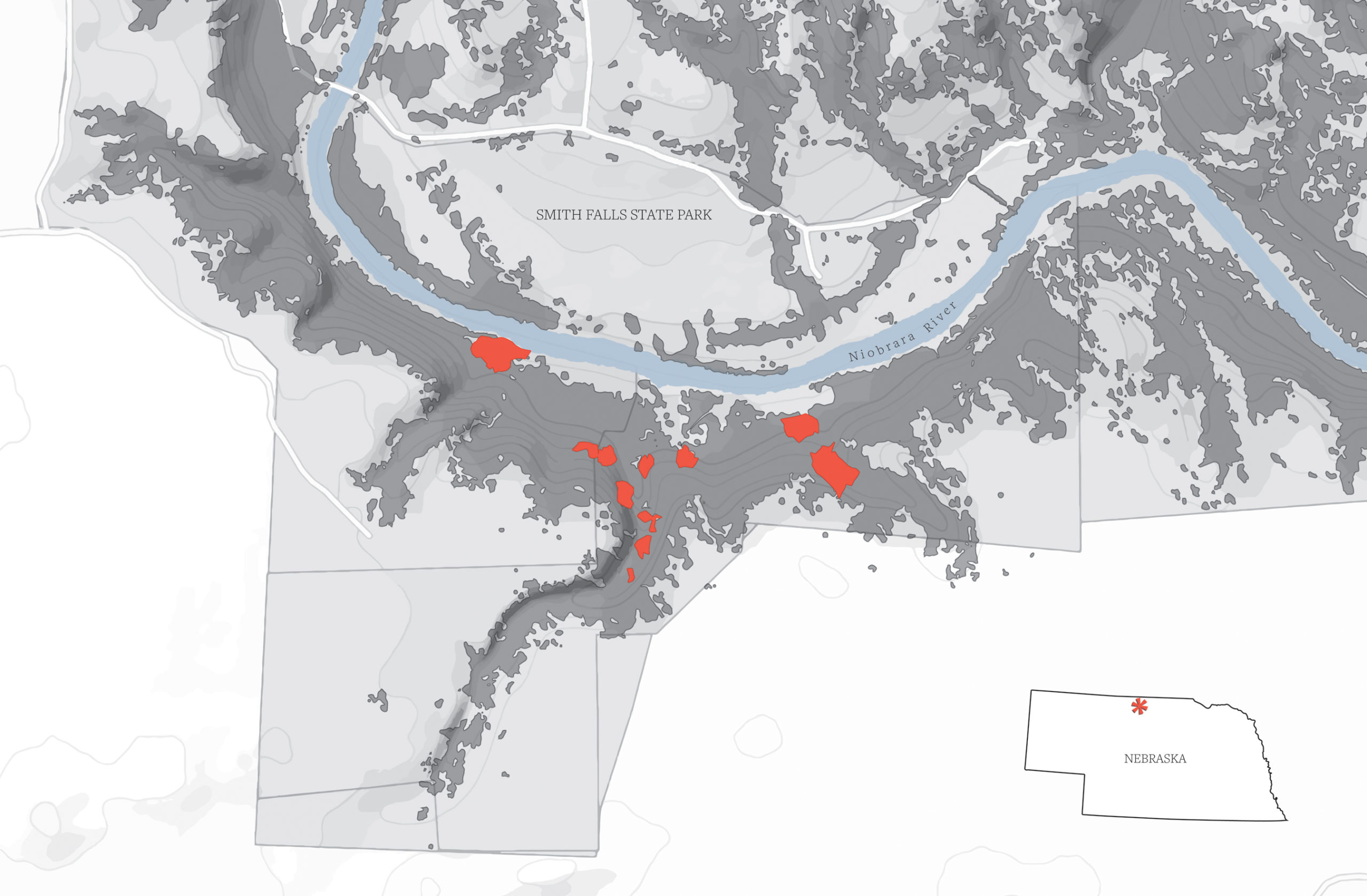
Aspen stands are vanishingly rare, if not altogether absent, in the Great Plains. Quaking aspens (Populus tremuloides) are nearly the sole representatives in the region but are restricted to a few tiny islands, usually growing in dense clumps around wetlands. These stands are relics, groups of trees left behind as the global climate warmed over the last ten thousand years. At the end of the Pleistocene Ice Age, aspen species, as with other cold-hardy trees, were probably quite common across this massive inland plain (Wright et al., 1985), but their ranges retracted as the climate became more arid. Stands of quaking aspen in central Canada and in the biotic highways of the Niobrara River Valley provide the only linkages between eastern and western populations for that species. My PhD advisor had secured funding from the National Parks Service—the Niobrara is a National Scenic River—to study one such stand, a rare, and therefore locally famous, collection of aspens centered around Smith Falls State Park, near Valentine, Nebraska. In 2013, we headed out for the first time to see these trees for ourselves.



When we began, it was immediately clear that the aspens at Smith Falls bear a great resemblance to quaking aspens. They have lovely white stems and dark green, heart-shaped, finely toothed leaves. They grow in clumped stands, indicating spread by rhizomes, with no singletons off on their own. We found these trees growing in a string of ten stands extending along the Niobrara River Valley, from Nature Conservancy holdings in the east to private property in the west. The stands in between these two locations are one of the crown jewels of Smith Falls State Park and are well known to its many visitors, who also come to raft the Niobrara River, camp out along its banks, and see the eponymous waterfalls. The aspen stands are confined to the cool, north-facing banks of the river, and they’re undeniably beautiful trees. But, as aspen fanatics, we agreed with what we’d already heard about them: they seemed somehow different than the quaking aspens so familiar to us in Minnesota. Their bark, though light-colored, seemed rather green, and their leaves were rather large, with fewer and larger teeth, compared to a typical quaking aspen. Indeed, these trees had long been held by local botanists and natural historians to be hybrids between locally rare quaking aspen and bigtooth aspen—a species whose eastern range edge is currently estimated to fall around 375 miles (600 kilometers) to the east, near Ames, Iowa.
We knew that quaking and bigtooth aspen hybridize naturally within their range— renowned forest ecologist Burton “Burt” Barnes’s seminal work documented many such stands in Michigan (1961)—but the thought of this hybrid having occurred naturally so far outside the distribution of one of its putative parental species struck me, at least, as somewhat scandalous. Aspen pollen is wind-distributed and can travel long distances, but there is no evidence of successful pollination events occurring when around 375 miles separate male and female plants. As such, I expected that the reputation of the Smith Falls aspens as hybrids was nothing more than understandable wishful thinking. After all, bark and leaf traits can be plastic, and a more parsimonious explanation of the unusual appearance of the Niobrara trees was that they were simply an unusual, isolated stand of quaking aspen. So, an important first step in our research would be to compare the genetics and morphology of the Smith Falls aspens to that of known quaking and bigtooth trees.
Our research group collected leaf samples, from which we could extract DNA, from all ten stands of Smith Falls aspens. We also dug up rhizomes to produce cloned suckers and planted these suckers in a common garden at a research station in Minnesota. There, we could perform experiments on them without harming the precious Niobrara trees. For the sake of comparison, we also drove all over the Midwest, collecting leaf and rhizome samples from quaking aspens in the Black Hills of South Dakota and the Sandhills of Nebraska, from bigtooth and quaking aspens along their western range edge in Minnesota and Iowa, and from both species within the interior of their distributions in Minnesota and Wisconsin.
Our assessment of the genetics and physiology of the aspens took place over two years at the University of Minnesota campus. We used microsatellite genotyping—the same technology that allows for DNA fingerprinting in humans—to understand which trees we had sampled were distinct individuals and which were clones. This work, as well as sequencing of parts of our sampled trees’ chloroplast genomes, was possible thanks to the full genome for black cottonwood (Populus trichocarpa), which was produced by an international team of dozens of biologists led by Oak Ridge National Laboratory’s Gerald Tuskan in 2006—the first whole genome project carried out for a tree species. In our physiological experiments, we pushed stems and leaves from our common garden trees to the brink. We measured them exhaustively, and we then dried them out and froze them to mimic climate change-induced drought and post-budbreak freezes. We also tracked their phenology—the timing of their leafing out and loss of leaves in our common garden.
An Ice Age relic
The results of our genetics work (Deacon et al., 2017) left me picking my jaw up from the lab bench. Our first finding struck an ominous tone: the Smith Falls aspens are shockingly undiverse. We are confident that, across all ten stands at the site, any given tree belongs to one of three genotypes. This means that three original seedlings produced through sex have given rise, through rhizome suckering, to all of the extant aspens in the area. We found genetically identical individuals growing on opposite sides of ravines and in stands separated by hundreds of meters. It appears, then, that these particular trees rely almost exclusively on asexual suckering for reproduction. More shocking still, at the nuclear level, the Smith Falls aspens shared genetic information with both quaking aspen and bigtooth aspen, confirming that they are a hybrid between these parents—a hybrid christened, appropriately enough, Populus × smithii. Evidence from chloroplast DNA suggested that bigtooth aspen, the species now not found until the middle of Iowa, was probably the maternal parent, with pollen coming from quaking aspen. Furthermore, patterns of genetic mixing we observed offer some support for the classification of these trees as F1 hybrids, meaning they are the first-generation offspring between two parents of different species, like mules.


Our study of leaves collected from common garden trees supported our finding that the Smith Falls aspens were in fact hybrids of quaking and bigtooth parents (Deacon et al., 2017). Use of a dichotomous key to distinguish between these species will often require inspection of the pubescence and margins of the leaves of the specimen in question. Quaking aspens tend to be glabrous with many small teeth on their leaf margins. Bigtooth aspens tend to be pubescent with fewer, larger teeth. Barnes’s work on Populus × smithii tells the same story. Though we documented many subtle differences between leaves of the two species in our systematic analysis, our findings contributed to the current consensus: pubescence and tooth number are the best way to tell them apart. And leaves from P. × smithii trees grown in Minnesota from Nebraska-collected rhizomes were perfectly intermediate between their putative parents in these two traits.
Taken together, these findings suggest the rather shocking story that, at some point when both bigtooth and quaking aspen were locally abundant in the Niobrara River Valley—probably between three and six thousand years ago—the two species hybridized. Both parents went locally extinct (and bigtooth vanished from the entire region), but their hybrid remained, reproducing vegetatively through rhizomes rather than through flowers and seeds. Despite considerable environmental change—the climate in this region was warming and drying long before our present human-induced bout of climate change—these aspens hung on, perhaps shrinking in their distribution, but not disappearing from one small stretch of the Niobrara River Valley. As such, these aspens are a true relic of a past climate and a unique genetic treasure of the region.
Specter of climate change
Our grant from the National Parks Service enabled us to go beyond determining the genetic identity of these trees. We also used our common garden to study their disturbing demographic decline. State managers and conservationists had noted that existing aspens looked stressed and that new trees either were not sprouting or were quickly consumed by deer before outgrowing their reach. We wanted to understand the vulnerability of the Smith Falls aspens to two forms of physiological stress likely to be concomitant with climate change. The first is straightforward: climate change in the region is likely to lead to more arid conditions, imposing drought stress on the vulnerable, mesic species of the Niobrara River Valley. The second is less so: because the forces that cause this warming do not necessarily prevent late-winter cold snaps, even if spring temperatures arrive earlier, plants can leaf out in response to an early spring, then get hit with a freeze after budbreak. Such post-budbreak freezing can range from damaging to catastrophic, potentially killing vulnerable tissues and leading to whole-plant death (Anderegg et al., 2015).
Our exploration of the aspens’ vulnerability to climate change, currently in review, resulted in some bad and some good news. Unsurprisingly, given aspens’ low level of drought tolerance, we found quaking, bigtooth, and hybrid aspens to be vulnerable to drought-induced cavitation, the formation of air bubbles in stem xylem. These bubbles, in quantity, disrupt waterflow in trees’ vascular systems, like holes in a straw. They can, ultimately, lead to total hydraulic failure and tree death. Quaking aspen was slightly more vulnerable to this type of drought damage than bigtooth aspen, and their hybrid was intermediate. More generally, the hybrid aspens shared some drought-tolerance traits with quaking aspen, others with bigtooth aspen, and, in other cases, was intermediate between the two. But all three taxa showed a limited capacity to resist the challenges likely to occur in a warming and drying climate.
The story surrounding post-budbreak freezing was simpler and rosier. We froze growing stems and leaves at temperatures equivalent to and lower than those that aspens in Nebraska are likely to experience during March and April storms. They were not substantially injured by this, suggesting that drought threatens aspens in the Niobrara River Valley more than late-winter cold snaps. Interestingly, in our measurements of spring phenology—the transition from dormancy to budbreak—we also found that the Smith Falls aspens were intermediate between quaking and bigooth aspen. Our findings echoed previous work showing that quaking aspens break bud about a week faster than bigtooths; fittingly, we observed that their hybrid offspring tended to leaf out in between the two parental species. Yet, compared to other regional conspecifics, all three groups of aspens generally leaf out around the same time.
An aspen in a juniper’s world
Though our determination that the Smith Falls aspens are in fact hybrids has proven fascinating and satisfying, thornier questions remain about their future. Locally, the Smith Falls aspens are being outcompeted by neighboring trees, especially eastern red cedars (Juniperus virginiana). Quaking and bigtooth aspens are tolerant of low-intensity fires but intolerant of shade. Fire exclusion has, therefore, been catastrophic for aspens, whether at Smith Falls or more broadly across the West. Because cedars are fire intolerant but drought tolerant, they have capitalized on our modern tendency to suppress and prevent fires. Locally, managers have fought this cedar encroachment by instigating prescribed burns, clearing cedars, and creating barriers to deer browsing on aspens. As a result, suckering is on the rise in some stands, producing a new cohort of healthy aspens (Robertson et al., 2018). Yet cedar removal and prescribed burns are expensive and will need to be repeated periodically to keep aspens abundant at Smith Falls.


Globally, climate change is also likely to reshape the distribution of aspens across North America. Nebraska is expected to experience climate warming in the decades to come, in tune with the current global commitment to three to four degrees of warming. And though rainfall in the region will likely remain stable in absolute quantity, rain and snow will fall more sporadically, producing longer and more frequent periods of drought. None of this is good news for aspens in the region. Given historical changes in the distribution of quaking and bigtooth aspen, observations colleagues and I have made suggest the Smith Falls aspens will encounter a greater risk of climate-induced extirpation in the Great Plains than they have faced since the end of the last Ice Age.
We might ask, then, what the future holds for the Smith Falls aspens and for other glacial relics in the region. The answer to this question depends on management. Stands of quaking aspen, paper birch (Betula papyrifera), and other drought-sensitive trees can probably be protected through active steps to shield them from direct climate stress and competition from more drought-adapted neighbors. In addition to prescribed burns, possible management practices include removal of competitors, use of exclosures to reduce grazing, targeted planting, and conservation of local groundwater. For species such as the aspens, which can be more easily propagated through rhizome cuttings than from seed, collection of rhizomes and propagation of suckers represents one pathway toward conservation of unusual, threatened germplasm. At present, private individuals and institutions may be able to assist with the migration of the Smith Falls aspens by purchasing commercially available nursery stock. Faller Landscape, in York, Nebraska, presently sells clonal trees produced through suckering from rhizomes collected at the site (marketed under the cultivar name ‘Ice Age’).2 More broadly, any efforts to stabilize the global climate, if our society is willing and able to undertake them, will also benefit Midwestern aspens, among many other species.
Hopeful monsters or dead ends?
Like other natural hybrids, the Smith Falls aspens have been heralded, at least locally, as uniquely adapted to their surroundings. It is tempting to assert that these trees, by virtue of their longevity, may illustrate one strategy for persistence in a warming, drying climate. Yet findings from our research do not support this narrative. Indeed, these hybrids are unique, and worthy of study insofar as they constitute an evolutionarily unusual relic from a past climate. But as noted above, these trees represent essentially three genetic individuals that have probably been cloning themselves for thousands of years. Though we have very recently received second-hand confirmation that the trees do flower in some years, we do not know the sex of each clone, or whether their flowers are fertile. Genetic evidence certainly suggests that no new trees have been born from seeds at the site for a very long time. And the Smith Falls aspens are beset by environmental challenges ranging from the hyperlocal to the global. Unfortunately, the Smith Falls aspens may simply have survived by demographic good fortune, constituting an evolutionary dead end, rather than a way forward.

Yet this is a rather pessimistic view, and, having grown rather fond of the aspens at Smith Falls during my time working with them, I think their conservation is justified. As unusual hybrids that have persisted despite millennia of climate change—whether by chance or due to some particular adaptation we did not measure—they represent a potential genetic resource. As such, they may be candidates for assisted migration: transplantation from their current, imperiled location to one that will be more appropriate in the coming centuries of a climate determined by human-induced changes. Propagation of these trees in cooler and wetter climates well within the current ranges of their parent species might allow them to flourish while also buying time for intentional propagation of second-generation hybrids through breeding with other Popolus × smithii or backcrossing.
Regardless, we would be wise to remember that the story of evolution in response to a warming climate has occurred many times in the history of life, and it often produces messy stories like that of the Smith Falls aspens. What appear to us to be non-adapted (or even maladapted) trees may hold the key to surviving and thriving in a future climate. After all, the common ancestor of modern aspens and willows had likely evolved during the earth’s last period of extreme warming, some fifty-five million years ago, when the tropics extended up to the North Pole (Manchester et al., 2006). Though the fate of quaking and bigtooth aspens and their hybrids is uncertain, the aspen lineage is likely to survive contemporary climate change. And so, whether hybrid aspens are best thought of as hopeful monsters or evolutionary dead ends is ultimately unknowable. But as an unlikely yet arguably successful hybrid myself, I’m inclined to give them the benefit of the doubt.
Acknowledgements
The original research reported in this article was conducted collaboratively at the University of Minnesota—Twin Cities with Jeannine Cavender-Bares, Nick Deacon, Anna Schweiger, and Isabella Armour. We were funded by National Park Service grant #191779 to Jeannine Cavender-Bares, Mark Dixon, and Molly Nepokroeff. Jeffrey Carstens, a horticulturist and curator of woody and herbaceous plants at the USDA’s North Central Regional Plant Introduction Station in Ames, Iowa, played an absolutely essential role in guiding our collections of Midwestern aspens, especially along the species’ range edges in Iowa. “Buffalo” Bruce McIntosh of the Nebraska Wildlife Federation and Joseph Zeleznik at North Dakota State University also provided essential help with collections in Nebraska and northern Minnesota, respectively. Three photographs in this article came from the video “Fighting for Survival: The Ancient Aspen of the Niobrara Valley,” produced by “Buffalo” Bruce McIntosh, Rod Jensen, and The Great Plains Motion Picture Company.
Notes
1 It should be noted that the chimaeras resulting from the almost unbelievable process of interspecies grafting (e.g. apples, pears, grapes, roses, citrus, mangos, stone fruit, and others, not to mention the introduction of certain porcine organs into human bodies) certainly give more integrated hybrids a run for their money in the realm of public interest.
2 Aspen enthusiasts can also purchase from them a very robust and aesthetically pleasing quaking aspen genotype, ‘NE Arb’, which was collected from a now-extinct stand elsewhere in Nebraska.
References
Anderegg, W. R. L., A. Flint, C. Y. Huang, L. Flint, J. A. Berry, F. W. Davis, J. S. Sperry, and C. B. Field. 2015. Tree mortality predicted from drought-induced vascular damage. Nature Geoscience, 8: 367–371.
Barnes, B. V. 1961. Hybrid aspens in the Lower Peninsula of Michigan. Rhodora, 63: 311– 324.
Bessey, C. E. 1887. A meeting-place for two floras. Bulletin of the Torrey Botanical Club, 14: 189–191.
Burger, W. C. 1975. The species concept in Quercus. Taxon, 24:45–50.
Deacon, N. J., J. J. Grossman, A. K. Schweiger, I. Armour, and J. Cavender-Bares. 2017. Genetic, morphological, and spectral characterization of relictual Niobrara River hybrid aspens. American Journal of Botany, 104: 1878–1890.
Goldschmidt, R. 1940. The material basis of evolution. New Haven, CT: Yale University Press.
Hamzeh, M., and S. Dayanandan. 2004. Phylogeny of Populus (Salicaceae) based on nucleotide sequences of chloroplast trnT-trnF region and nuclear rDNA. American Journal of Botany, 91: 1398–1408.
Kaul, R. B., G. E. Kantak, and S. P. Churchill. 1988. The Niobrara River Valley, a postglacial migration corridor and refugium of forest plants and animals in the grasslands of central North America. The Botanical Review, 54: 44–81.
Manchester, S. R., W. S. Judd, and B. Handley. 2006. Foliage and fruits of early poplars (Salicaceae: Populus) from the Eocene of Utah, Colorado, and Wyoming. International Journal of Plant Science, 167: 897–908.
Robertson, J. M., A. R. Cahlander-Mooers, and M. D. Dixon. 2018. Effects of management treatments on regeneration of a geographically disjunct, relictual hybrid aspen (Populus × smithii) population in the central Great Plains, USA. Environmental Management, 62: 906–914.
Tuskan, G. A, S. Difazio, S. Jansson, J. Bohlmann, I. Grigoriev, U. Hellsten,… D. Rokhsar. 2006. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science, 313: 1596–604.
Citation: Grossman, J. J. 2019. The hybrid mystique. Arnoldia, 76(3): 2–13.
Wright, H. E., J. C. Almendinger, and J. Grüger. 1985. Pollen diagram from the Nebraska Sandhills and the age of the dunes. Quaternary Research, 24: 115–120.
Wu, G. A., J. Terol, V. Ibanez, A. López-García, E. Pérez-Román, C. Borredá,… M. Talon. 2018. Genomics of the origin and evolution of Citrus. Nature, 554: 311–316.
The maps in this article were created using ESRI, USGS, USFS, NGA, NASA, CGIAR, N Robinson, NCEAS, NLS, OS, NMA, Geodatastyrelsen, Rijkswaterstaat, GSA, Geoland, FEMA, Intermap and the GIS user community.
Jake J. Grossman is a Putnam Fellow at the Arnold Arboretum.