
When you sip an aromatic Riesling with dinner in September, the day may feel noticeably shorter than it did a month before. While fall officially starts with the autumnal equinox, which occurs between September 21 and 24 from year to year, day length continuously decreases after the summer solstice and will continue through the arrival of winter. From August to November, in fact, the days get shorter by almost three minutes per day in Boston. Plants, including the common grapevine (Vitis vinifera), pay close attention to day length from the moment leaves and flowers unfurl in the spring until fruit ripens and leaves drop. Although this internal clock may seem difficult to conceptualize—more esoteric than flavor and mouthfeel, or even pests and diseases—understanding the process by which plants enter and exit dormancy, and how they survive in between, is critical, especially as winegrowers (and other agricultural producers) adapt their production to withstand a changing climate.

As grapevines grow, they form buds in the leaf axils. Within these buds, about ten nodes are pre-formed—leaf primordia (baby leaves) and inflorescence primordia (baby flowers). These buds are formed in preparation for the following growing season. In that way, wines produced in 2018 actually started as microscopic inflorescences in the summer of 2017, which overwintered inside buds, formed flowers that bloomed in the spring of 2018, and became fruit that was harvested in the late summer and fall, effectively spending over a year developing on the plant alone. This process doesn’t happen only in Vitis vinifera, the species native to the Mediterranean from which most of our wines are produced. Wild grape species like the riverbank grape (V. riparia), which is native to North America, and gloryvine grape (V. coignetiae), which ranges from eastern Russia to Japan, go through the same process, as do the majority of temperate perennial plants.
Unlike migratory birds that avoid winter by flying south, plants are stationary. Therefore, plants must endure low temperatures and other unsuitable growing conditions that come with the winter. In these conditions, they lower their metabolism and enter dormancy, much like hibernation for animals. Plants, however, have evolved to start the preparation for winter much before damaging temperatures arrive, without relying on year-to-year weather patterns. Leaves register the decreasing day length (or technically the increasing nighttime), which provides a signal for buds to enter dormancy. Plants then start changing color to create the beautiful spectacle of fall foliage. In this process, grapes relocate nutrients from the leaves into storage tissue in the woody vines, ready to be recycled the following year. Once plants lose their leaves, they no longer track the day length. Instead, grapevines know when to start growing in the spring by tracking how long they have been cold.
Much like a person setting an alarm to have eight hours of sleep, buds count the number of hours spent in what are called chilling temperatures, between 32 and 50°F (0 and 10°C). Just as different people need different amounts of sleep to be productive, each grapevine species, and even different genotypes within a single species, have different chill requirements before they are able to come out of dormancy. Once they have accumulated enough chilling hours, buds are able to better respond to warm temperatures and produce spring growth. The chilling requirement is associated with the region where a species originated: species from lower latitudes are accustomed to low chill accumulation because of short winters, while species from higher latitudes have a high chill requirement because of longer winters. Given that no chill accumulation occurs below 32°F (0°C), however, plants growing in cold continental climates, like Minnesota, or much higher latitudes, like northern Canada, experience low chill accumulation. Species from these places, therefore, also tend to have a low chill requirement.
Chill requirements prevent plants from resuming growth during midwinter warm spells, which could cause death of both flowers and leaves upon the return of cold weather. The mechanisms that make buds wait for the appropriate and consistent warm weather in the spring are still largely unknown, but solving this mystery is important. As temperatures continue to increase due to climate change, the amount of chilling that plants experience in different areas is changing: lower latitudes are experiencing fewer hours with chilling temperatures, whereas the opposite is true for higher latitudes, like Boston and upstate New York. This trend in northern areas may seem a little counterintuitive, but time that was previously spent in below-freezing temperatures is now rising into the chilling range, above freezing but below 50°F (10°C), causing chill accumulation to rise.


When plants fail to accumulate the necessary chilling requirement, they have erratic, reduced, and delayed budbreak. In vineyards and orchards, this means reduced yields. The same is true with native forests, where flowering and corresponding seed production drops. Moreover, shifting phenology could detrimentally impact overlap between flowering and the activity of pollinators for a given species, or there may be competition due to overlapping flowering for species that were previously staggered. In areas where excessive chill is expected, more responsive plants may break buds during midwinter warm spells, when they previously would have known to wait. In agricultural settings, new cultivars and adaptive management practices can help overcome these effects in the short term. For forests, however, climate change is happening faster than floristic composition can change. Researchers at the University of Alberta, in Canada, have estimated that forest species are already 80 miles south of their optimal climate niche, and this figure is expected to increase to 190 miles in the 2020s, beyond recorded rates of natural forest migration.
Dormancy Fundamentals
Vitis is a compelling genus to study dormancy because of its distribution from tropical to subarctic regions. Even the common grapevine (V. vinifera) alone is capable of growing within a wide band of climates, with the majority of production in the Northern Hemisphere spanning latitudes between San Diego and Vancouver (30 to 50° north). In the Southern Hemisphere, the band is even wider, stretching from northern Argentina to southern New Zealand (20 to 50° south), with some production occurring as close to the equator as northeastern Brazil (9° south). Moreover, many of the most popular cultivars like V. vinifera ‘Pinot Noir’ and ‘Chardonnay’ are present in almost all of these areas, which demonstrates the remarkable plasticity of the species. Also, many varieties have been cultivated for a very long time. The first records for ‘Riesling’ date from the fifteenth century—before Columbus arrived in the Americas—and impressively, seeds of ‘Rkatsiteli’ were found in clay vessels dating to 3,000 BC. ‘Rkatsiteli’ is still grown in Georgia, the eastern European country where the archaeological discoveries occurred, and limited production can be found in the Finger Lakes wine region of New York. This historical continuity provides us with a wealth of records from different regions, providing both temporal and geographic context for understanding the basic requirements for dormancy of grapevines.
Measuring the chilling requirement of different grapevine varieties can be very simple, and in fact, similar techniques can be used to study dormancy in most deciduous perennial plants. Cuttings with dormant buds that have experienced different amounts of chill are placed in a warm environment (72°F or 22°C) with sixteen hours of daylight. The chilled buds can either be collected from the field in regular intervals throughout the winter, or collected in late fall or early winter and placed in a cold chamber where they’ll be removed after certain doses of chilling have been provided. The number of days taken for budbreak decreases for buds that have experienced additional chilling. When at least 50 percent of the buds have expanded within twenty-one days of being put into warm conditions, the buds are considered to have fulfilled dormancy requirements, and have now moved into a different phase in the dormancy cycle.


Dormancy can be divided into three phases. Paradormancy occurs during the growing season due to factors that arise outside the bud tissue, typically from plant hormone concentrations. For example, hormones produced in the shoot tip prevent lateral growth too close to the tip. This phenomenon, known as apical dominance, dictates the general conical shape of spruce trees (Picea), for instance, because the uppermost buds are more suppressed than the lower buds. Because grapevines are pruned, paradomancy is minimized, allowing lateral branching throughout the growing season. Endodormancy occurs when factors within the bud prevent growth. This phenomenon is triggered by decreasing day length and temperatures in the autumn. Endodormant buds resist growth, even when taken from the outside and placed in a warmer environment. Ecodormancy, as the prefix eco hints, occurs when environmental factors prevent the resumption of growth. Buds transition from endo- to ecodormancy through chill accumulation. Essentially, plants clock how long the winter has been, and this tells them whether they should grow when exposed to warm temperatures (spring has arrived), or if they should remain dormant (midwinter warm spell). Once the buds are ecodormant, they will only remain in a dormant state so long as temperatures remain low.
The necessity for chilling is one of the major factors that determines the distribution of vineyards in subtropical and temperate climates. In regions where insufficient chilling occurs naturally, grapevines and other fruit tree species require extra help to transition from endo- to ecodormancy. Wine growers can apply artificial stressors, such as agrochemicals (e.g. hydrogen cyanide), natural compounds (e.g. garlic extract), or even heat from mobile ovens, to jolt endodormant buds into an unnatural ecodormant state, much like a blaring alarm clock. Yet even though these methods can bypass chilling requirements, none are capable of stimulating budbreak as synchronously as natural processes.
In addition to dormancy, plant tissues must have mechanisms to cope with below-freezing temperatures. Leaves and other green tissues are generally very sensitive to freezing, due in part to their high water content. This is why deciduous plants lose their leaves in the fall. In the case of grapevines, buds survive the winter through a process called supercooling. The precise mechanisms that contribute to differences in supercooling ability are exceptionally complex and not well characterized, but at the most basic level, supercooling allows water to be cooled below 32°F (0°C) and remain liquid. In fact, given the right conditions, water can remain liquid to temperatures around -40°F (-40°C), although once frozen, the water will only melt at 32°F (0°C). You might have observed this phenomenon if you have ever left a water bottle in your car overnight when the temperature dropped below freezing. When you grab the bottle the following morning, the water may still be liquid, but when you shake the bottle, ice will immediately start forming. Supercooling does not necessitate altering the concentration of sugars or other metabolites—the antifreeze method used in a car engine—rather, with grapevines and many temperate fruit species, physical barriers to ice formation seem to play major roles in the supercooling process.
Supercooling provides variable levels of cold hardiness for grapevine buds throughout the winter. If the outside temperature drops below the current level of cold hardiness, ice will form inside the bud tissues, killing the tiny leaves and flowers beneath the bud scales. This means that the threshold temperature for ice formation changes throughout late fall, winter, and early spring. As temperatures begin to dip below freezing in the late fall, grapevine buds slowly begin to gain cold hardiness, gradually increasing their ability to survive freeze events. The buds always must remain ahead of the environmental temperature, which is very important, and in New York, the first freeze event of the season rarely occurs before grapevines have gone dormant due to this process of acclimation. Under artificial conditions in growth chambers, we have found that if temperatures are held or oscillated around 27°F (-3°C), dormant buds can acclimate to survive temperatures as low as -4°F (-20°C). But the process isn’t instantaneous and may take up to two weeks.
Dormancy Danger
Understanding the mechanics of dormancy matters because winter is dangerous for grapevines and other perennial plants, and when it comes to agricultural production, predictable harvests are paramount. Depending on the climate of different grape-growing regions, the most perilous time of winter can differ. In New York, slow temperature decline during the fall helps grapevines fully prepare for winter. In contrast, in the Pacific Northwest—an increasingly respected wine region, known for producing European-inspired vintages—the coldest days of the year are often in early December, and rapid temperature changes can zap buds before the acclimation process is finished.
Wine growers in New York don’t escape unscathed; instead problems arise due to midwinter cold temperatures. Because the vast majority of commercial grape cultivars have been selected from Vitis vinifera, the only grapevine species native to the Mediterranean and therefore adapted for hot summers and mild winters, these cultivars can only survive to a maximum cold temperature of around -4 to -13°F (-20 to -25°C). This temperature range is not unheard of in upstate New York, often causing at least partial bud dieback each winter—the reason vineyards in this region are mostly located around the Finger Lakes and on the shores of the Great Lakes. About sixty wild grape species can be found across North America and Asia, and most have greater maximum cold hardiness than the common grapevine. For example, the fox grape (V. labrusca)—the North American species from which ‘Concord’ grapes were selected—can endure temperatures around -22 to -31°F (-30 to -35°C), or even lower. Amur grape (V. amurensis), which has a broad distribution throughout eastern Asia, may be capable of surviving even lower levels. But while these species seem to promise natural cold hardiness for breeding efforts, our ability to tap into this genetic variation is relatively limited, given that pure water can only supercool to about -40°F (-40°C). Thus, winemakers in regions where winter temperatures drop below this level must be satisfied with importing grapes.
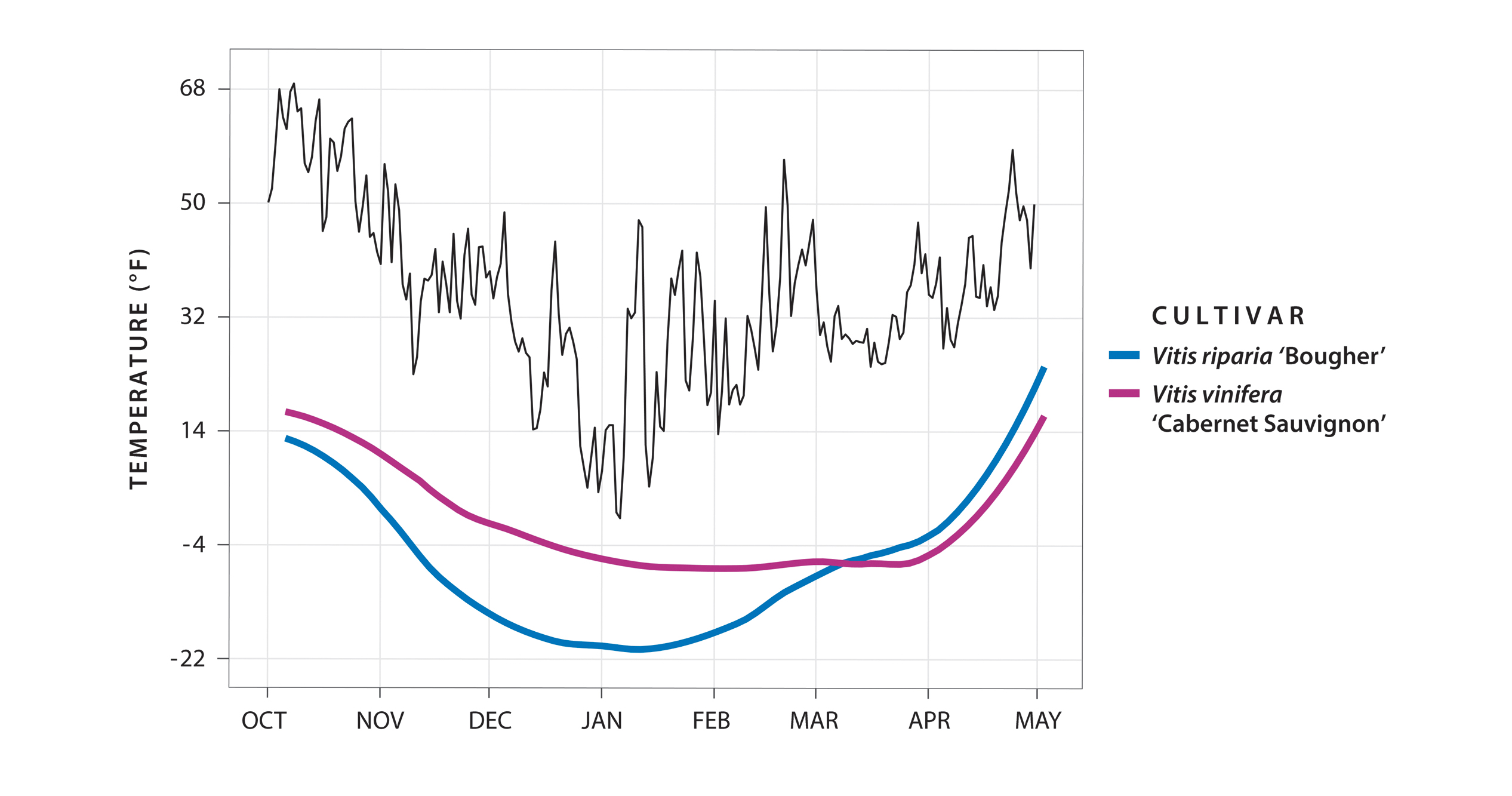
If grapevine buds survive acclimation and midwinter temperatures, the next major event occurs when the buds start losing their cold hardiness as spring temperatures warm. We call this process deacclimation. This is another time of great risk, and climate change promises to make this transition even more unpredictable. Climate models suggest that polar vortex events will become more common in late winter and early spring, catching buds during deacclimation and resulting in lethal damage. Luckily, different species deacclimate at different speeds. The riverbank grape (Vitis riparia) tends to be much faster than the common grapevine (V. vinifera), for instance, while the southern wild grapevine (V. cinerea) seems to be much slower. This difference in northern and southern species probably indicates natural adaptations as a result of their respective winter climates. As a northern species, V. riparia probably evolved a fast deacclimation rate to take advantage of the shorter summers, while V. vinifera and V. cinerea, each more adapted to milder winters and hotter summers, lack the adaptive need to race toward growth. In this way, wild grapevines provide us with the natural adaptive differences needed to learn about cold hardiness and dormancy, and also with the breeding material needed to meet the challenges of a changing climate.
Climate change has already reduced the amount of winter chill accumulation in most traditional wine regions. Bordeaux, the largest winegrowing area in France, now receives about 75 percent of the chilling it had in the mid-1970s. Our latest research has demonstrated that the speed of early spring development is dependent on chill accumulation, and that we can measure this speed based on deacclimation. So to understand the implications of current climate trends in regions like Bordeaux, more detail must be added to our analogy of dormancy as a night of sleep. When you sleep only one to two hours and get up, it feels like you have not rested at all. Once you hit three hours, every additional hour of sleep feels like a great improvement—think about the difference between four and five hours—although after seven hours, each additional hour provides less energy improvement. We were able to measure a similar phenomenon with the grapevine buds. With minimal chill accumulation, the buds lost their hardiness very slowly, but once they accumulated about eight hundred hours, there was a sharp increase in how responsive they were to warm temperatures. After about twelve hundred hours, however, there was little improvement to responsiveness—the equivalent of surpassing seven hours of sleep. In this sense, the transition between endo- and ecodormacy is gradual, not a clear dichotomy between states.

What does this mean for viticulture? Despite the reduced chill accumulation in Bordeaux, mentioned above, the region still receives plenty of chilling for Vitis vinifera cultivars. As a result, the buds and vines are usually ready to begin growing as soon as spring temperatures warm. In 2017, however, unseasonal warmth in April caused early budbreak as plants were very responsive due to high chill accumulation. A subsequent frost occurred in late April and caused extensive damage to vines, reducing the crop by 40 percent compared to the previous year, resulting in one of the lowest yields in the past thirty years. Thus, the future for wine production is complicated from the standpoint of dormancy, cold hardiness, and sustainable viticulture. Climate models predict less chill in warm regions in the future, leading to a need for different cultivars or the use agrochemicals to force vines. In cooler regions, increased chilling temperatures between 32 and 50°F (0 and 10°C) may lead to earlier budbreak for current cultivars, which could be especially detrimental given the increasingly erratic patterns of late winter cold events.
Production of wine is not the main concern with a changing climate, given that food production and broader ecosystem stability are at risk. While our research has focused on grapevines, we expect that similar behavior would be seen with many other horticultural and forest species. Plant distribution is governed by temperature, and these are generally predicted to increase in the future. This means the optimal zones for many plants will move towards the poles—especially if they require chilling. Agricultural production can more readily adapt with new cultivars and species, but forests may need a hand if we want to continue seeing the current diversity available. Minimum temperatures are the most limiting factor for plant distribution, so as the world gets warmer, it is perhaps a little ironic that increasing our understanding of how plants respond to cold may be key to predicting how they will survive in the future.
Additional reading list
Cook, B. I., and Wolkovich, E. M. 2016. Climate change decouples drought from early wine grape harvests in France. Nature Climate Change, 6(7): 715–720.
Gray, L. K., and Hamann, A. 2013. Tracking suitable habitat for tree populations under climate change in western North America. Climatic Change, 117(1-2): 289–303.
Gu, L., Hanson, P. J., Post, W. M., Kaiser, D. P., Yang, B., Nemani, R., and Meyers, T. 2008. The 2007 eastern US spring freeze: Increased cold damage in a warming world?. AIBS Bulletin, 58(3): 253–262.
Hannah, L., Roehrdanz, P. R., Ikegami, M., Shepard, A. V., Shaw, M. R., Tabor, G., and Hijmans, R. J. 2013. Climate change, wine, and conservation. Proceedings of the National Academy of Sciences, 110(17): 6907–6912.
Citation: Kovaleski, A., and Londo, J. 2018. Dormant Vines, Future Wines. Arnoldia, 76(2): 10–17.
Kovaleski, A. P., Reisch, B. I., and Londo J. P. 2018. Deacclimation kinetics as a quantitative phenotype for delineating the dormancy transition and thermal efficiency for budbreak in Vitis species. AoB PLANTS, 10(5): ply066.
Londo, J. P., and Kovaleski A. P. 2017. Characterization of wild North American grapevine cold hardiness using differential thermal analysis. American Journal of Enology and Viticulture, 68: 203–212.
Al Kovaleski is a postdoctoral researcher at the United States Department of Agriculture’s research station in Geneva, New York. He completed his doctorate at Cornell University in 2018.
Jason Londo is a research geneticist with the United States Department of Agriculture’s Grape Genetics Unit, based at the New York State Agricultural Experiment Station in Geneva, New York. He is an adjunct associate professor at Cornell University’s School of Integrative Plant Science.