
“The American chestnut once comprised 25% or more of the Native Eastern Hardwood Forest.” American Scientist (1988)
“Chestnut was perhaps the most widespread and abundant species in the Eastern United States since the last glaciation.” USDA Forest Service Southern Research Station General Technical Report General Technical Report SRS-173 (2013)
Before the turn of the century, the eastern half of the United States was dominated by the American chestnut.” American Chestnut Research and Restoration Project, SUNY College of Environmental Science and Forestry (2013)
Along with the bison and the passenger pigeon, the American chestnut forms an iconic triumvirate of the grandeur of the American wilderness and the devastation that human activity wrought upon it over the past three centuries. Just as the bison was the preeminent large mammal on the continent and the passenger pigeon the most abundant bird, so is chestnut often described as having dominated the eastern forest (or across its geographic range) prior to its destruction by an introduced Asian chestnut blight.
By all accounts chestnut was a magnificent and invaluable tree. It was among the fastest growing, tallest, and widest-trunked trees in the eastern United States. The strength, straight grain, and decay resistance of its wood made it ideal for framing, finished lumber, and fencing, and its regular production of nuts provided abundant food for native and European peoples, domestic livestock, and diverse wildlife. But was it really the dominant tree in the eastern forest?
Dominant species, in the words of forest ecologist E. Lucy Braun, are “those trees of the canopy, or superior arboreal layer, which numerically predominate.” Given American chestnut’s purported prior dominance in the eastern deciduous forest, we would expect the tree to have ranged widely across the East relative to other common tree species and to occupy a superior place in written accounts by early naturalists and explorers, early land survey records, forest surveys of the early twentieth century, and the paleoecological record. In fact, these sources reveal a very different story.

Accounts by Early Explorers and Naturalists
Accounts by foresters about chestnut’s abundance at the turn of the twentieth century have been widely cited in the scientific and popular literature as evidence of the tree’s former dominance. Descriptions of chestnut by naturalists and explorers at the time of European settlement, on the other hand, are rarely cited. Early written records must be used with caution, given that they were often written by non-botanists and provide a potentially biased assessment of previous forest conditions (Whitney 1994). Nonetheless, these descriptions—particularly if they correspond with other available lines of evidence—provide valuable eyewitness accounts of eastern forests prior to their widespread modification by European settlement. Below are selected quotations that reference chestnut and other species by some of the more important early explorers and naturalists in the Eastern United States.
John Smith, New England coast (early 1600s): “Oke [oak], is the chiefe wood, of which there is great difference in regard of the soil where it groweth; fir, pine, walnut, chestnut, birch, ash, elm …, and many other sorts.”
(Smith 1616)
Colonel William Byrd, Virginia (1737): “chestnut trees grow very tall and thick, mostly, however, in mountainous regions and high land …”
(Bolgiano and Novak 2007)
William Bartram, northern Alabama–Mississippi border (late eighteenth century): “[we entered] a vast open forest which continued for above seventy miles … without any considerable variation … the forests consist chiefly of Oak, Hiccory, Ash, Sour Gum, Sweet Gum, Beech, Mulberry, Scarlet maple, Black Walnut, Dogwood, Aesculus pavia, Prunus indica, Ptelea, and an abundance of chestnut on the hills, with Pinus taeda and Pinus lutea.”
(Bartram 1976)
Although these accounts represent only a very small sample of early observations, they offer some general patterns that are reinforced by many others not reported here, specifically that chestnut appears to have had a relatively restricted niche (mountainous) rather than being generally abundant throughout the landscape, and to have been secondary in importance to oaks (Quercus).
The Biogeography of Chestnut
The eastern deciduous forest spans approximately 926,000 square miles in North America, covering 13 entire states and substantial portions of 10 others from Maine to Minnesota and south to Texas and Georgia. This vast area is broadly united by a cover of deciduous or mixed deciduous-coniferous forest, but otherwise is far from uniform. Five climatic regions, twelve geomorphic regions, and five soil regions define this broad area. Climate, landforms, and proximity to the coast determine the frequency and type of natural disturbances (e.g., tornadoes, hurricanes, fires, ice storms) that influence a particular region, as well as the distribution and abundance of human populations and their disturbances such as tree cutting, agriculture, and the removal and introduction of wildlife. The physical environment and its associated natural and human disturbances, in turn, shape the vegetation.




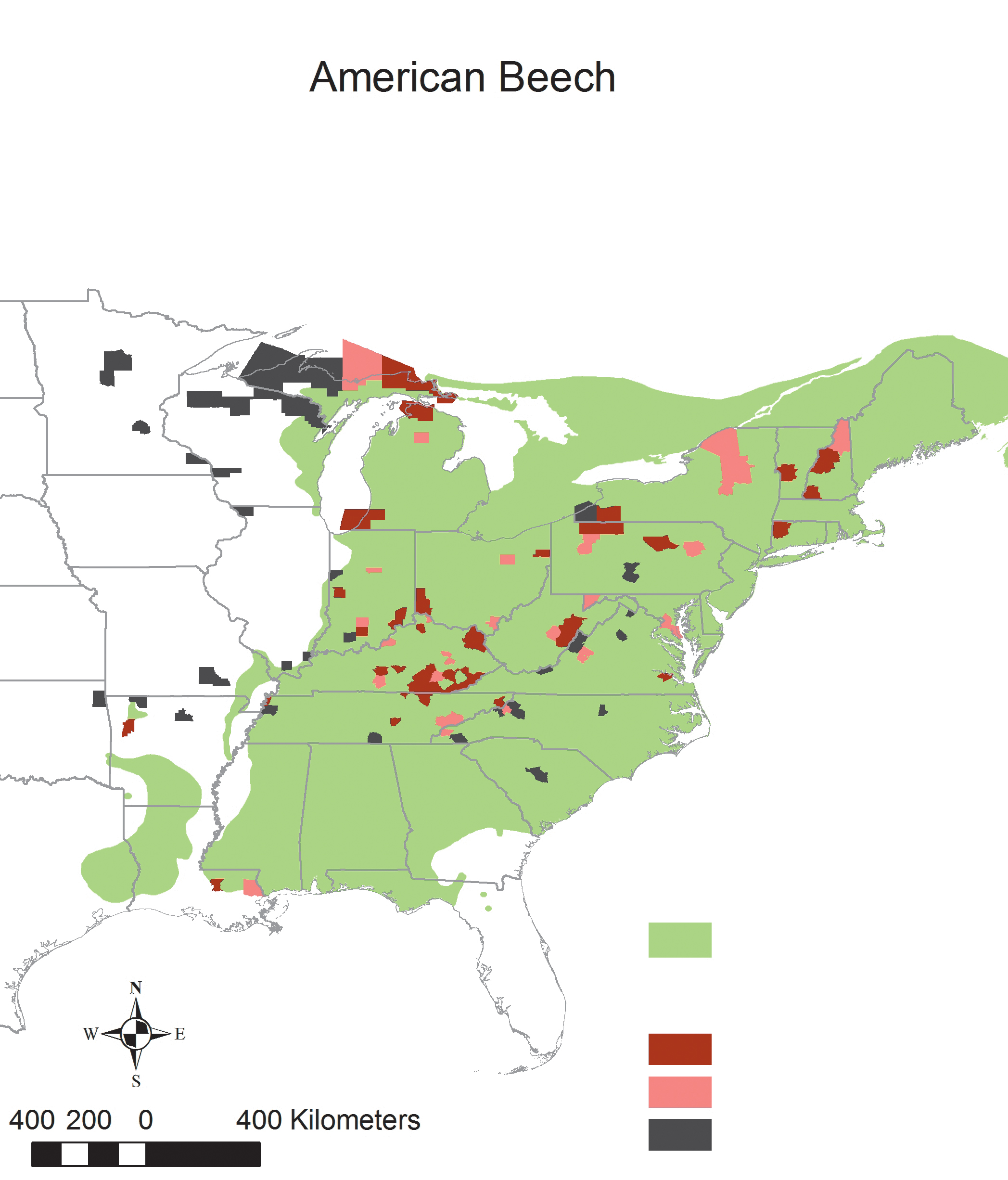
For a tree species to dominate an area as broad and diverse as the eastern forest it needs to be an ecological generalist. Relative to other common species like white oak (Quercus alba), American beech (Fagus grandifolia), red maple (Acer rubrum) and sugar maple (Acer saccharum), chestnut had limited ecological amplitude. Chestnut has high water requirements relative to oaks and is restricted to moderate climates. Hence, it grew predominantly—as the early explorers noted—in sloping topography, particularly on moist, well-drained lower slopes and on some rocky ridges. Chestnut generally fared poorly on sandy coastal plains and outwash soils, clayey soils, saturated wetland soils, or calcium-rich sites. Much of the southeastern coast of the United States is dominated by sandy soils and therefore lacked chestnut altogether. Large areas of the midwestern section of the eastern forest have calcium-rich soils and relatively low rainfall and were thus also unsuitable for chestnut. In northern New England, northern New York, and upper Michigan, extremely cold winters were largely prohibitive to chestnut, which is susceptible to cold and frost damage. In sum, chestnut ranged across only about 309,000 square miles of eastern North America in the early twentieth century—about one-third of the Eastern forest. In contrast, sugar maple, red maple, white oak, red oak (Quercus rubra), American beech, and American basswood (Tilia americana) all have geographic ranges that exceeded chestnut’s by at least a factor of three (Little 1971).
Witness Trees
Early land surveys conducted at the time of European settlement frequently utilized trees, known as witness trees, as corner posts and reference points, and surveyors often recorded each tree to genus or species. Compiled across counties, states, and regions, witness trees offer a formidable inventory of the forest composition that greeted the first European settlers. Early land survey data reveal that chestnut was far less abundant at the time of European settlement than the oft-quoted 25% of the forest. A recent paper by Jonathan Thompson, Charles Cogbill, and colleagues compiled witness tree data from over 700 townships from nine states in the northeastern United States. Their results show that chestnut comprised a mere 3% of trees in the region and never exceeded 25% of trees in a single town. In contrast, beech comprised 22% of trees across the region; oaks, predominantly white oak, 17.5%; and hemlock 11%.
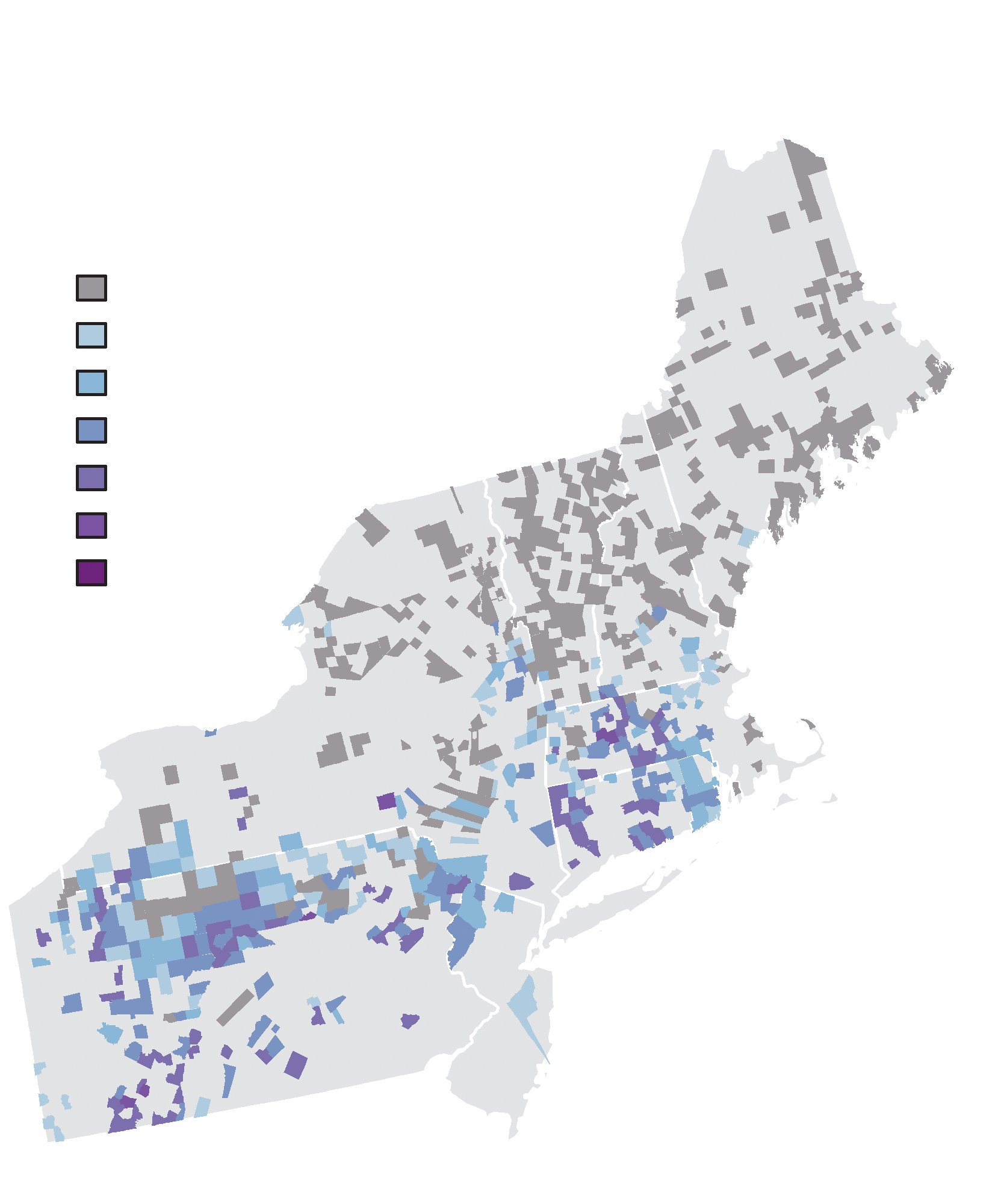


Two decades ago, forest historian Gordon Whitney compiled maps of tree species abundance from land survey data across the midwestern United States. Data from about 100 counties or townships across eight states of the upper Midwest reveal that chestnut was never the dominant tree, comprising 5 to 15% of trees in a small section of Ohio and 0 to 4% of trees in the rest of the region. In contrast, beech and especially white oak were frequently the dominant tree, often comprising 25 to 65% of all trees. Limited early land survey data from the southern regions of the eastern forest also portray chestnut as a secondary species. Chest nut was the first-ranked species in only one of 15 locations, whereas white oak was the first-ranked tree in five of 15 locations (see Table on facing page).
Early Twentieth Century Forest Surveys
E. Lucy Braun conducted and compiled extensive forest surveys and observations across 120 counties of the eastern forest in the early twentieth century. Her data were predominantly gathered from “original” forests and thus fill in gaps in the witness tree studies, particularly in regions such as the Cumberland Mountains of Kentucky and the Blue Ridge Mountains of North Carolina and Tennessee. Although Braun acknowledged her unequal coverage of different regions, her work remains by far the most comprehensive assessment of the eastern deciduous forest, including American chestnut’s abundance, at the time of the chestnut blight. Her surveys and data tables reveal that chestnut was a tree of surprisingly limited dominance. Chestnut was dominant (the most abundant canopy tree) in at least one survey in only 15 of the 120 counties (12.5%) sampled by Braun and others. Sugar maple, white oak, and hemlock were all dominant species in over 20% of the counties sampled, and beech was a dominant tree in over 40% of the counties sampled. In fact, Braun’s data suggest that chestnut was not even the most abundant tree within its own geographic range: beech was a dominant species in at least one survey in almost half (48%) of the counties sampled in chestnut’s range, whereas chestnut was a dominant tree in less than a quarter (23%) of the counties sampled.


American chestnut was spectacularly abundant in some locations. On north slopes in Joyce Kilmer Memorial Forest in North Carolina, for instance, it comprised over 83% of the canopy trees, and on the slopes of Salt Pond Mountain in western Virginia, it made up 56 to 85% of the canopy trees (Braun 1950). Chestnut could also grow to enormous size. In a forest in Central Kentucky, Braun wrote that chestnuts, which comprised 22% of the canopy trees, were “by far the largest trees, about 5 feet d.b.h. (diameter at breast height).” But chestnut was far from the only tree to achieve such local dominance; beech, hemlock, sugar maple and white oak all achieved comparable abundances in other stand locations. In 1876, forester A. R. Crandall wrote the following in eastern Kentucky: “white oak has a wider range and greater development in numbers than any other species. In size it ranks with the largest of the hardwood trees …”
The Rise of Nineteenth Century Logging and Chestnut
In its destructiveness and lack of legal control, nineteenth century commercial logging was similar to the unrestricted hunting that decimated the passenger pigeon and the bison. However, in an ironic twist to the story of American chestnut, this particular act of exploitation actually promoted chestnut to dominance in parts of its range where it hadn’t been dominant before. Chestnut’s remarkable ability to sprout vigorously from cut stumps, including those of large diameter and advanced age, made it better adapted to intensive logging than any other hardwood tree including oaks. As the early Connecticut foresters Hawley and Hawes (1912) wrote, “this sprouting capacity of the species is its strongest characteristic and the one by which with each successive cutting it gains in the struggle for existence with the rival inmates of the woodlot.” Interestingly, chestnut’s sprouting capacity was much more prominent in the Northeast than in the southern parts of chestnut’s range. In heavily cutover forests of northern New Jersey and southern New England, chestnut increased from 5 to 15% of the forest during the early colonial period to an estimated 50% of the standing timber in Connecticut. Because Braun focused on “original” forests in her surveys, she largely avoided surveying the cutover southern New England region so her data probably underestimate chestnut’s abundance in the Northeast. But it’s important to remember that southern New England represents a small fraction of chestnut’s range and the eastern forest overall.


The Last to Arrive: Chestnut Since the Last Ice Age
Fossil pollen records in the Eastern forest enable reconstruction of vegetation communities and tree species that have dominated forests over the past 15,000 to 50,000 years. In formerly glaciated areas such as the Northeast, pollen records provide a chronological record of recolonization of forest vegetation after glacial melt some 15,000 to 20,000 years BP (before present). In southern New England, ash (Fraxinus), birch (Betula), ironwood (both Ostrya and Carpinus, whose pollens are indistinguishable from each other), and oak arrived first, followed by maples; deciduous forests replaced coniferous forests about 9,000 years BP. Beech arrived about 8,000 years BP, and hickory about 6,000 years BP. Not until about 2,000 years BP does chestnut pollen appear in the sediment record, earning chestnut the distinction of being the last major tree species to recolonize the region after deglaciation (Davis 1983). When chestnut finally does appear in the sediment record, it generally doesn’t exceed about 4 to 7% of the pollen types across the region with the exception of one record in northwestern Connecticut where it reaches 18 to 19% (Paillet 1991, Oswald et al. 2007). In contrast, oak pollen consistently comprises 40 to 60% of the pollen and beech 5 to 20%. Interestingly, chestnut does achieve great dominance (40 to 70%) at the stand scale in a few local New England pollen records (Foster et al. 1992, 2002), exemplifying the importance of spatial scale when considering the abundance of this species.
What accounts for chestnut’s late arrival to New England? One possible reason is that the climate of the Northeast throughout much of the Holocene was too dry for chestnut. Other researchers have posited a lack of favorable well-drained germination sites in southern New England after deglaciation, or too much lime in the soil that took millennia to leach away. Chestnut is also self-sterile unlike many other trees that are self-fertile, and thus the chances of establishing new populations were much lower for this tree. Whether dispersal or environmentally limited, it is clear that chestnut was poorly adapted to recolonizing the deglaciated Northeast compared to other hardwood trees.


Chestnut had a much longer history in the unglaciated Southeast. Chestnut pollen appears in the pollen record as early as 16,000 years BP in Tennessee (Davis 1983). Although a few records show chestnut to be dominant or co-dominant with oaks during the Holocene in the North Carolina and Tennessee mountains, most of the records from the southern and central Appalachians analyzed by William Watts, Paul and Hazel Delcourt, and others reveal oaks to be dominant over chestnut. Still, comparisons between oak and chestnut pollen abundance should be undertaken with caution. Oak pollen grains are indistinguishable among species, and many are therefore combined into a single category of “oak” pollen. Chestnut, on the other hand, is the only species in its genus in the Northeast and is one of two species (the other is dwarf chinkapin, Castanea pumila) in the central and southern Appalachians. Oak pollen is wind dispersed and therefore is generally produced in larger quantities than is chestnut pollen, which is partially dispersed by insects. Hence, chestnut pollen is generally underrepresented in the pollen record, relative to oaks. Still, chestnut’s relatively minor status in the pollen record is consistent with its secondary status in the witness tree data and in accounts by early settlers. In addition, chestnut’s great abundance (40 to 45%) in a few southern Appalachian pollen records analyzed by the Delcourts and stand-level records from Massachusetts are consistent with twentieth century forest surveys in which chestnut achieved great dominance in some landscapes and topographic positions, but generally not at broader scales.
Concluding Thoughts
American chestnut was once a common tree species throughout its Appalachian Mountain range and a dominant species in parts of its central and southern range (primarily the oak-chestnut forest region). However, prior to European settlement, it was less dominant than white oak and beech and far less widespread than most other major tree species. With increasing timber harvesting in the nineteenth and early twentieth centuries, chestnut’s dominance increased in the northern part of its range in heavily cut-over forestland. Still, the tree remained absent from fully two-thirds of the eastern forest, precluding it from ever being the dominant tree of this biome.

Revealing the truth about American chestnut’s relatively limited place in the Eastern forest does not diminish the grandeur of this great tree, its historical importance to cultures of the central and southern Appalachians, and the great tragedy of its demise. Chestnut remains the flagship example of the potential dangers posed by introduced pathogens in our native forests. But we should be careful not to let a great tragedy and impassioned restoration efforts trump the available data when discussing the history of this tree.
A Killer Arrives
Chestnut blight (Cryphonectria parasitica) was first discovered in 1904 in a stand of American chestnuts (Castanea dentata) in New York’s Bronx Zoological Park, perhaps arriving on imported nursery stock of Castanea crenata from Japan. Subsequent investigation determined that the blight arrived in the late nineteenth century, as evidence suggested that American chestnuts on Long Island had been infected as early as 1893. The effects of the blight were immediate and devastating, often killing mature trees in 2 to 3 years. By 1906, the blight was detected in New Jersey, Maryland, and Virginia and continued to spread rapidly, reaching Pennsylvania in 1908 and North Carolina by 1923. All government efforts to contain or eradicate the blight failed, and ceased entirely by 1915. By the early 1940s the destruction of the American chestnut throughout its 300,000-square-mile range was complete.

The blight spreads by wind-borne fungal spores that invade the tree through cracks or injuries in the bark, killing the cambium and eventually girdling the tree. The roots generally survive the blight, however, and continue to produce sprouts that are eventually killed again before reaching reproductive age. In effect, the chestnut blight converted a once towering overstory tree into an understory shrub.
Figure | Early Land Survey Data
Dominant tree species and corresponding abundance and rank of American chestnut at the time of European settlement identified from early land survey data in the southeastern United States. Adapted from Abrams (2003).
| Location | Dominant Tree Species and Abundance (%) | Chestnut Abundance (%) | Chestnut Rank | Reference |
| Eastern West Virginia – Ridge and Valley | White oak (33) | 5 | 5 | Abrams and McCay 1996 |
| Eastern West Virginia – Allegheny Mts. | Beech (13) | 6 | 8 | Abrams and McCay 1996 |
| Southern West Virginia | White oak (24) | 12 | 2 | Abrams et al. 1995 |
| Northern Virginia | White oak (49) | 0 | NA | Orwig and Abrams 1994 |
| Southwestern Virginia | Red oak (25) | 9 | 3 | McCormick and Platt 1980 |
| Western Virginia | White oak (26) | 5 | 5 | Stephenson et al. 1992 |
| Central Georgia | Pine, mostly loblolly and shortleaf (27) Post oak (18) | 2 | 9 | Cowell 1995 |
| Northeastern Georgia | Pine (26) American chestnut (20) | 20 | 1 | Bratton and Meier 1998 |
| Southcentral Tennessee | Post Oak (11) | 2 | 11 | DeSelm 1994 |
| Northern Florida | Magnolia (21) | 0 | NA | Delcourt and Delcourt 1977 |
| Southeastern Texas | Pine, mostly longleaf (25) | 0 | NA | Schafale and Harcombe 1983 |
| Southeastern Louisiana | Magnolia (13) | 0 | NA | Delcourt and Delcourt 1974 |
| Northeastern Louisiana | Pine, longleaf, shortleaf, and loblolly (24) White oak (11) | 0 | NA | Delcourt 1976 |
| Eastern Alabama | Pine, 7 species (44) Post oak (12) | 2 | 9 | Black et al. 2002 |
| Southern Arkansas | Black oak (18) | 0 | NA | Bragg 2003 |
Spatial Scale
Spatial scale refers to the size or extent of the area under consideration. A stand is a relatively small area of forest that is spatially continuous in structure and composition and is exposed to similar soil and climatic conditions. In paleoecology the size of the catch basin (e.g., lake, pond, swamp, or small hollow) determines the distance from which pollen in the sediments originates. Sediments from a small forest hollow will contain pollen from vegetation growing predominantly in the immediate stand (a “stand scale” investigation), whereas sediments from a large lake are dominated by pollen from the broader landscape up to 20 miles away.
References
Abrams, M. D. 2003. Where has all the white oak gone? Bioscience 53: 927–939.
Abrams, M. D., D. A. Orwig, and T. E. DeMeo. 1995. Dendroecological analysis of successional dynamics for a presettlement-origin white pine– mixed oak forest in the southern Appalachians, USA. Journal of Ecology 83: 123–133.
Abrams, M. D., D. M. McCay 1996. Vegetation-site relationships of witness trees (1780–1856) in the presettlement forests of eastern West Virginia. Canadian Journal of Forest Research 26: 217–224.
American chestnut research and restoration project. 2014. https://www.esf.edu/chestnut/background.htm
Bartram, W. A. 1976. Travels and Other Writings. Princeton: Princeton University Press.
Black, B. A., H. T. Foster, and M. D. Abrams. 2002. Combining environmentally dependent and independent analyses of witness tree data in east-central Alabama. Canadian Journal of Forest Research 32: 2060–2075.
Bolgiano, C. and G. Novak. 2007. Mighty Giants: an American Chestnut Anthology. American Chestnut Foundation, Bennington, VT.
Bragg, D. C. 2003. Natural presettlement features of the Ashley County, Arkansas area. The American Midland Naturalist 149:1–20.
Bratton, S. P. and A. J. Meier. 1998. The recent vegetation disturbance history of the Chattooga River Watershed. Castanea 63: 372–381.
Braun, E .L. 1950. Deciduous Forests of Eastern North America. Caldwell, New Jersey: The Blackburn Press.
Burnham, C. R. 1988. The restoration of the American chestnut. American Scientist 76: 478–487.
Cowell, C. M. 1995. Presettlement Piedmont forests: patterns of composition and disturbance in central Georgia. Annals of the Association of American Geographers 85: 65–83.
Davis, M. B. 1983. Quaternary history of deciduous forests of eastern North America and Europe. Annals of the Missouri Botanical Garden 70: 550–563.
Delcourt, H. R. 1976. Presettlement vegetation of the north of Red River Land District, Louisiana. Castanea 41: 122–139.
Delcourt, H. R. 1979. Late quaternary vegetation history of the eastern Highland Rim and adjacent Cumberland Plateau of Tennessee. Ecological Monographs 49: 255–280.
Delcourt, H. R and P. A. Delcourt. 1974. Primeval magnolia-holly-beech climax in Louisiana. Ecology 55: 638–644.
Delcourt, H. R. and P. A. Delcourt. 1977. Presettlement magnolia-beech climax of the Gulf coastal plain: quantitative evidence from the Apalachicola River bluffs, north-central Florida. Ecology 58: 1085–1093.
Delcourt, P. A. and H. R. Delcourt. 1998. The influence of prehistoric human set fires on oak-chestnut forests in the southern Appalachians. Castanea 63: 337–345.
Delcourt, P. A., H. R. Delcourt, P. A. Cridlebaugh, and J. Chapman. 1986. Holocene ethnobotanical and paleoecological record of human impact on vegetation in the Little Tennessee River Valley, Tennessee. Quaternary Research 25: 330–349.
DeSelm, H. R. 1994. Vegetation results from an 1807 land survey from southern middle Tennessee. Castanea 59: 51–68.
Foster, D. R., T. Zebryk, P. Schoonmaker, and A. Lezberg. 1992. Post-settlement history of human land-use and vegetation dynamics of a Tsuga canadensis (hemlock) woodlot in Central New England. Journal of Ecology 80: 773–786.
Foster, D. R., S. Clayden, D. A. Orwig, B. Hall, and S. Barry. 2002. Oak, chestnut, and fire: climatic and cultural controls of long-term forest dynamics in New England, USA. Journal of Biogeography 29:1359–1379.
Hawley, R. C. and A. F. Hawes. 1912. Forestry in New England. New York: John Wiley and Sons.
Little, E. L., Jr. 1971. Atlas of United States trees, volume 1, Conifers and Important Hardwoods: Misc. Pub. 1146. Washington, D.C.: U.S. Department of Agriculture. 9 p., 200 maps
McCormick, J. F. and R. B. Platt. 1980. Recovery of an Appalachian forest following the chestnut blight or Catherine Keever—you were right! American Midland Naturalist 104: 264–273.
Orwig, D. A. and M. D. Abrams. 1994. Land-use history (1720–1992), composition, and dynamics of oakpine forests within the Piedmont and Coastal Plain of northern Virginia. Canadian Journal of Forest Research 24: 1216–1225.
Oswald, W. W., E. K. Faison, and D. R. Foster. 2007. Postglacial changes in spatial patterns of vegetation across southern New England. Journal of Biogeography 34: 900–913.
Paillet, F. L. 1991. Relationship between pollen frequency in moss polsters and forest composition in a naturalized stand of American chestnut: implications for paleoenvironmental interpretation. Bulletin of the Torrey Botanical Club. 118: 432–443.
Russell, E. W. B. 1987. Pre-blight distribution of Castanea dentata (Marsh.) Borkh. Bulletin. Torrey Botanical Club 114: 183–190.
Shuman B., P. Newby, Y. Huang, and T. Webb III. 2004. Evidence for the close climatic control of New England vegetation history. Ecology 85: 1297–1310.
Smith, J. 1616. A Description of New England. Electronic Texts in American Studies Libraries at University of Nebraska–Lincoln.
Stephenson, S. L., H. S. Adams, and M. L. Lipford. 1992. The impacts of human activities on the upland forests of western Virginia. Virginia Journal of Science 43: 121–131.
Citation: Faison, E. K., and Foster, D. R. 2014. Did American chestnut really dominate the eastern forest? Arnoldia, 72(2): 18–32.
Thompson, J. R, D. N. Carpenter, C. V. Cogbill, and D. R. Foster. 2013. Four centuries of change in northeastern United States forests. PLoS ONE 8: e72540.
Wang, G. G., B. O. Knapp, S. C. Clark, and B. T. Mudder. 2013. The Silvics of Castanea dentata (Marsh.) Borkh., American chestnut, Fagaceae (Beech Family) Gen. Tech. Rep. SRS-GTR-173. Asheville, NC: U.S. Department of Agriculture Forest Service, Southern Research Station. 18 p.
Watts, W. A. 1979. Late quaternary vegetation of central Appalachia and the New Jersey coastal plain. Ecological Monographs 49: 427–469.
Watts, W. A. 1980. The late quaternary vegetation history of the southeastern United States. Annual Review of Ecology and Systematics 11: 387–409.
Whitney, G. G. 1994. From Coastal Wilderness to Fruited Plain: A History of Environmental Change in Temperate North America from 1500 to the Present. New York: Cambridge University Press.
Edward K. Faison is Ecologist at Highstead in Redding, Connecticut, and David R. Foster is Director of Harvard Forest, Harvard University.