When I began as a field researcher on the island of Borneo, in 1957, little was known about the distribution of the inland rainforests. The forests are incredibly diverse and are dominated by large overstory trees in the dipterocarp family (Dipterocarpaceae), which often tower more than two hundred feet above the forest floor. While variation in the forests was evident, it appeared as chaotic and random as the colored specks in a children’s kaleidoscope. Odoardo Beccari, a Florentine botanist who spent two years in northern Borneo in the 1860s, had provided the first confirmation that specialized lowland habitats, including peat swamps and sandy exposures, bear distinct forest types, but neither he nor his successors until my time had recognized any correlation between habitat characteristics and forest structure on the yellow-to-red tropical soils that characterize much of the inlands.

Yet as I tramped along Bukit Biang—a long ridge in eastern Brunei—I was surprised to see dipterocarp species I had come to know on the sandy coastal hills of western Brunei, and I began to sense that these forests were divided into two distinct communities—one on sandy soils, the other on loams. These communities would reappear in different localities as I extended my explorations throughout Brunei. I came to anticipate the flora by the distinctive sounds of the cicadas that inhabited each and by the smells of the forest, which I later recognized all over Borneo and even Peninsular Malaysia—the mellow fruitfulness and fermentation from the loams or the resinous aroma from the peaty humus covering the sandy soils. Those forest smells returned to me decades later, after I had assumed the directorship of the Arnold Arboretum and first trekked into the loamy bottomlands in the Connecticut River Valley and the sandy pine barrens of New Jersey and Cape Cod. It was only then that I came to understand that these habitat patterns in the forested landscapes of mild, moist climates are universal. Yet, in Brunei, I began to sense that individual species within these rainforest communities were often more highly habitat specific than I had ever seen in temperate forests.
After twenty-eight intensive months of fieldwork, camping, and longhouse life in Brunei, I made the case that differentiating between these inland forest types—known collectively as mixed dipterocarp forests (or MDFs)—could have important implications for timber inventories and silviculture. At that time, timber was only cut for local use in northwestern Borneo, although research towards sustainable harvesting was advanced in Peninsular Malaysia. Even there, as elsewhere in the tropics, distinct types of lowland MDFs had yet to be defined. The forestry department gave me clearance and funding to lay out sets of plots to test my hypothesis. For foresters, understanding the distribution of these tree communities could guide sustainable harvesting practices. But knowledge of tree species preferences and distributions would also provide the means of mapping biodiversity, locating centers of richness and endemism, and identifying and demarcating priorities for conservation—a first for the tropics.
~
Work on the research plots commenced in 1959. But first, I briefly returned to England for my own wedding. My wife, Mary, was to become the perfect companion for a life of jungle exploration. She had been born and spent her first years in Sri Lanka, where her family had been in trade and tea for over a century. She would join my hectic field life at once, and we only had a few days in town before departing for a long stay in the hulu (or upriver country). I wanted to document and compare the two main forms of MDFs that I had recognized in my explorations. I decided to compare two seemingly contrasting sites—one on the sandy coastal hills of Andulau, in western Brunei, and the other on steep clay-loam ridges near Kuala Belalong, in eastern Brunei. I sensed that topography was an important feature, causing local variation in the form, composition, and possibly growth rates of the forests, so at each site, I planned for fifty one-acre plots that covered the complete topographic spectrum (including ridgetops, slopes, valley bottoms, and riverbanks). Each one-acre plot represented a homogeneous topography, and the number and size of the plots were visually estimated to be representative of the forest variation at each site. This method had already become a standard procedure for ecologists studying temperate grasslands, but it was a first for biodiverse tropical forests and, indeed, for any forests known to me. All trees above one-foot circumference were to be documented. We measured the trunk girth and estimated the height for each. We also identified each as morphologically different species by means of fallen leaves and local Iban names. But I rarely knew the scientific name for these visually distinct trees.



Life eventually settled down to calm and peace at out first location, Kuala Belalong. It was one of the most beautiful places in which we ever camped, with the rush of water over the rapids in our ears as we lived and slept. While I scrambled daily up the muddy slopes, locating plot positions, surveying, and initiating documentation with the teams, Mary would take her typewriter to the shingle beach, preparing herbarium labels and editing field notes. She saw the wildlife that I rarely or never experienced: a pair of small-clawed otters (Aonyx cinerea) that came to join her, squeaking and gamboling in the shallow water nearby; the extraordinary and terrifying pack of Bornean beared pigs (Sus barbatus), thundering headlong down the steep hillside opposite, then splashing across a rapid upriver, on migration in search of fruiting trees; and a macaque (Macaca fascicularis) who discovered our food store and made off with some delicacies.
Our work in Andulau began the following year, in 1960, and was carried out smoothly, given our growing experience and the gentle landscape. I continued to conduct multi-week collecting expeditions until I was satisfied that the diversity of landscapes and forests in Brunei had been examined.
~
When we returned to the University of Cambridge, after the 1960 field season, I brought back two suitcases of fallen leaves and twenty notebooks—all needed to complete my doctoral dissertation. I had presorted the leaves into nearly eight hundred morphospecies, which were recognizable entities that were mostly (dipterocarps excepted) lacking a formal scientific name and therefore named using indigenous nomenclature. They were to be named with the help of taxonomists at the University of Leiden and the Royal Botanic Gardens, Kew, supported wherever possible by our herbarium-quality specimens in flower or fruit. This groundwork provided the basis for a Checklist of Brunei Trees, which I co-authored with Hasan bin Pukol, the ascendant curator of the Brunei Herbarium, who had been a mentor on tree identification from the time he joined our team. He was an experienced informant on traditional plant uses and became a good friend who involved Mary and me in his family activities, including marriages and births (and he even got us invited to a royal circumcision).
Our combined efforts eventually provided accurate names to support the baseline data from our one hundred plots: some twenty-five thousand individuals representing nearly seven hundred species. I had no idea how I could sort and compare the plots using this elephantine data set! But luck came my way, for a few months after my arrival in Cambridge, the annual meeting of the British Ecological Society was convened there. I attended, where I was recommended to confess my rash achievements to Peter Greig-Smith, a professor at the University College of North Wales, Bangor, who was one of the leaders of a new science known as quantitative plant community ecology. Greig-Smith, a modest if somewhat austere academic, asked whether I had seen the recent paper, published in Ecological Monographs, on the woodlands bordering the tallgrass prairies of northern Wisconsin. Authors Roger Bray and John Curtis of the University of Wisconsin were to save my career. They had devised a method of relating their forests to one another, and to their climate and soils, by comparing plots according to the presence and abundance of each species present. This general method is known as ordination, and elements of their novel approach are still used today.
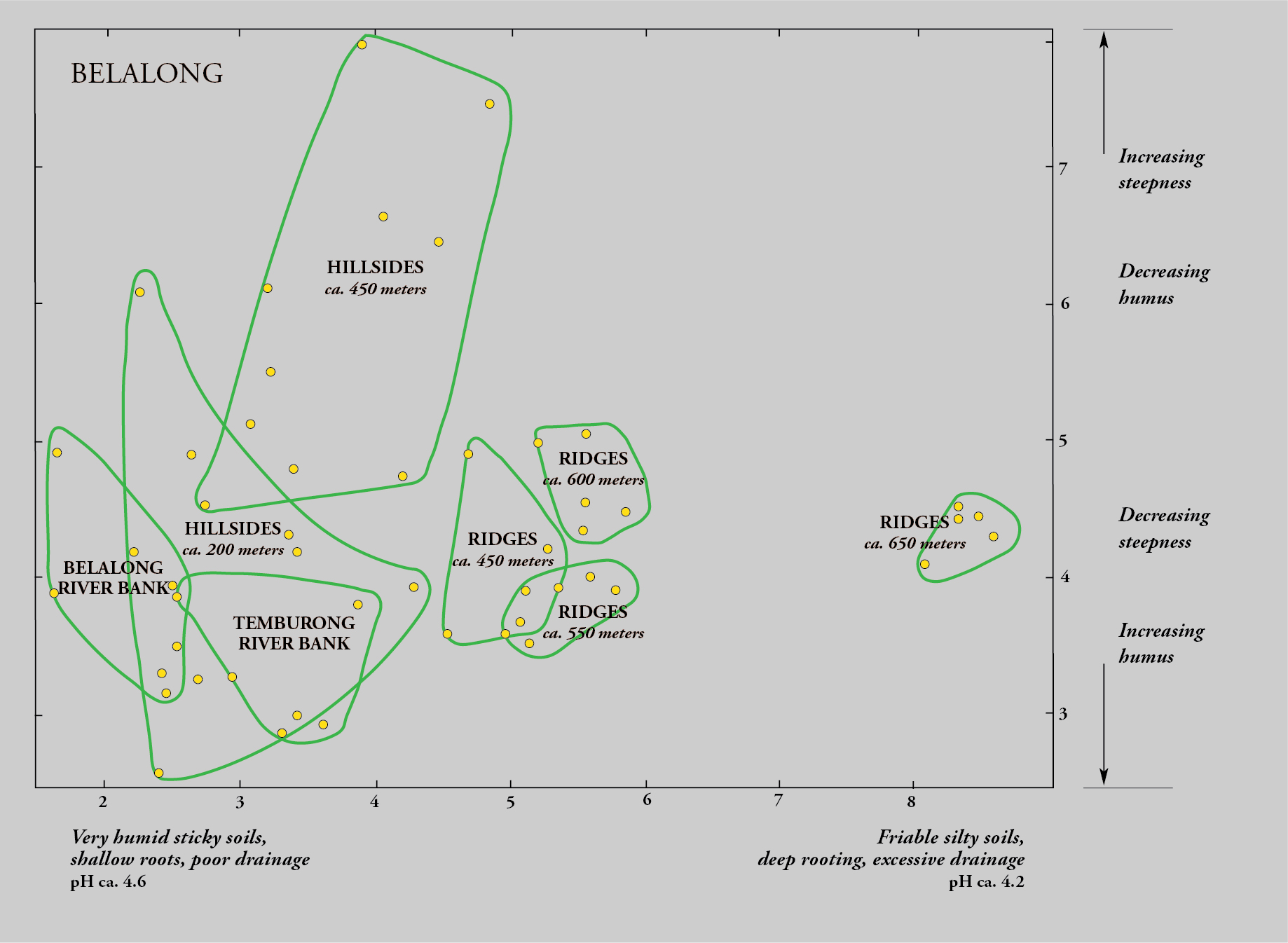
Examination of the data from Brunei indicated that the tree flora at Belalong was markedly different from that at Andulau, with only a third of species common to both sets of plots, so I decided to ordinate the two sets separately. I set to work, entering the plot data and calculating the matrix of similarity indices. I toiled night and day, for more than three weeks, using a bulky hand calculator, the Swedish Facit. The day of reckoning arrived when I started to place the plots, as dots, using a simple geometric technique for ordination, in two dimensions so that the distance between them related to their floristic similarity. To my amazement, a recognizable pattern gradually emerged for the fifty plots at each site, much like, in those days, how a photographic print would emerge on paper set in hypo solution. The patterns confirmed intuitions gained from field experience, with the tree flora tied intimately with geology and topography.
Although we hadn’t recensused the plots to show change over time, the initial results were clear. The species composition indicated that the dipterocarp species on clay loams, and particularly the lower slopes and undulating land, were predominantly light hardwoods that grow relatively fast. These species might yield a timber crop within a half century. But the sandy soils of Andulau, and also the shallow loams along the sharp Belalong ridges, were dominated by heavy dipterocarps and other hardwoods whose growth rates were known from other research to be much slower, implying that more complex management would be needed to sustain selective felling, with growth cycles exceeding a century.
The ordinations demonstrated, for the first time, that hyperdiverse tropical lowland forests were as floristically variable and habitat-specific as temperate broadleaf forests; indeed, the individual species showed a degree of habitat specificity only found in temperate forests in specialized habitats such as limestone crags. It is the tree species, through their chemical and physical interactions with other forms of life, that directly or indirectly mediate all biodiversity. This knowledge provided a breakthrough, and I knew that the work needed to be expanded—both geographically, to see whether the same trends would be observed elsewhere on Borneo, and temporally, to observe changes in the forest structure over time.
~
Mary and I, now with a young family, returned to Borneo in 1962. I resumed my explorations, this time as forest botanist in the adjacent state of Sarawak (which became part of Malaysia in 1963), where we eventually spent five years. It was a perfect place to bring up our three children. Soon we were taking them to the woods where we, looking up into the canopy for signs of flowering, would be nudged by them, looking down with a detail to be observed from their stature alone, alerting us to a nest of giant ants, or a huge millipede, or a leech swaying encouragingly.



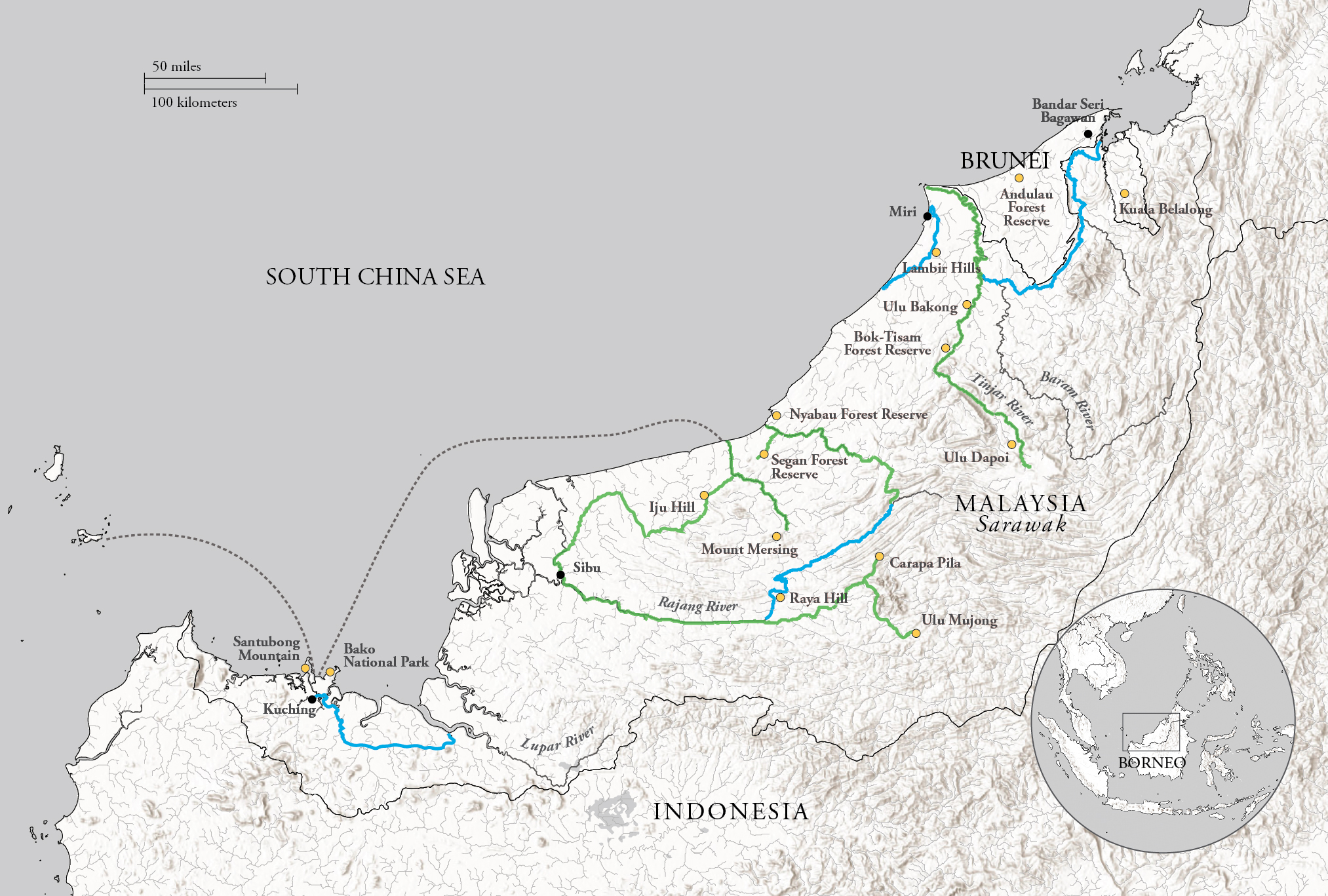
Sarawak is slightly larger than New York State and therefore twenty times the area of Brunei. This presented both a challenge and an opportunity. I adopted a plan whereby, as in Brunei, I would undertake one major botanical exploration each year, while making periodic short forays when time allowed. Although Sarawak and Brunei share a dominant sedimentary geology of sandstones, shales, and clays, Sarawak also supports tantalizing habitats on isolated pockets of limestone karst and volcanic rocks. I further suspected that the major waterways—the Baram, Rajang and Lupar Rivers—could provide evidence of separate diversification, even speciation, in the lowland forests isolated on either side of their extensive floodplains. The major expeditions provided opportunities to set up plot clusters in a diverse set of habitats and geographical locations. As in Brunei, plot results would add to knowledge needed to understand patterns of timber supply and quality. We would also have opportunities to initiate recensusing of permanent plots, conducted on five-year intervals. This would allow us to test predictions of growth rates and management protocols inferred from the static data gathered in Brunei.
Our first year of plot surveys started with a crisis: The December-to-February northeast monsoon came in with a fury not matched in recent history. Kuching, in the west, where we all lived, received continuous rain rattling on our roofs for more than two weeks at a time. Over one-third of the state was underwater, but miraculously, no lives were lost thanks to the army with their inflatable rivercraft. But it also provided an opportunity to establish permanent plots on landslide locations, uniquely allowing us to monitor forest regeneration from scratch.
Afterwards, these journeys became routine, socially as well as botanically. For the first few hours upriver, travelling in a dugout canoe known as a prahu, we would pass through mangrove forests to the first Malay fisherman’s town where we would check in at the government office, buy food, and learn whether our advance party had succeeded in attracting local Dayaks—the indigenous people of Borneo—who were familiar with the terrain and who would also provide boats. We would then proceed to the forest, often spending nights in riverside longhouses along the way. Each longhouse is essentially a village of wooden rowhouses, on stilts and under one roof, entered by ascending a notched log (like a ladder) at either end. The shared roof shelters a gallery on whose floor all social activities flourished.
These stayovers were always hilarious, if somewhat raucous, experiences. As the chosen house approached, our team of local field assistants would begin combing and oiling their hair, smartening up and adding perfume; when we turned the corner and saw the longhouse for the first time, as often as not there would be a group of young women, having heard the noise of our outboards, who would have descended to the landing to wave and shout encouragement. Conversations on these boat rides were always alive with joking and good-natured braggadocio.
At first, the riverside vegetation would consist of a mix of cultivated trees—indigenous mango and durian species, rambutans, coconut palms, and native and Cavendish bananas—and the indigenous species of the floodplains. But things would change as soon as the current quickened and the first rocky banks were exposed. A distinct flora appeared below the flood line: miniature palms, aroids, ferns, and a diversity of shrubs and coarse herbs, known as rheophytes, which are adapted to periodic immersion and sweeping floodwaters—a community rich in rare species awaiting collection. Overhead, trees that had gained traction on the rocks leaned precariously over the narrowing water, their branches dripping with epiphytic ferns, orchids, and even rhododendrons (Rhododendron sect. Vireya). We observed trees like Dipterocarpus oblongifolius, which produces bright-pink winged fruit that hang like Christmas decorations, and the fragrant babai (Saraca declinata), a leguminous species, which produces decorative yellowish-orange flowers on its trunk.

At this point in the upriver journey, those with boating skill came to the fore, led by the outboard operator and the individual stationed on the prow with a long suar—a fending pole—who was known as the jagar luan (or prow-guard). The most crucial moment was always at the head of a rapid when the prahu, lifted during its ascent, would drop onto the calm water as we entered a lagoon. The stern would lift, and with it the propeller. All hands took to the poles for, if we failed to heave the baggage-filled prahu across, we could lose control, fall back headlong, probably sideways, and lose everything to the torrent. I experienced this seven times, but thanks to willing and experienced hands, we never lost our precious specimens, notebooks, or soil samples, although a camera could get a dousing. I would watch and admire the skill shown. But upriver travel was punishing to the outboard motor: what would start as a shiny new Evinrude with fancy hood would, on occasion, end up as an unprotected swirling stick, topped with a greasy bareboned engine—still miraculously spinning!


We aimed to establish plots representing the full range of yellow-red tropical soils and the MDFs that dominate the lowland Bornean landscape. In addition to recording representative forest profiles along transects, at sites of uniform geology, I had suspected that much of the floristic variation was influenced by soil fertility, perhaps individual nutrient ions, as in temperate forest communities. Soil analytical laboratories hadn’t been available in Brunei, but in Sarawak, facilities were available. So, this time around, we sampled soils: at the surface, where organic duff was concentrated, and at a standard depth of 30 inches (75 centimeters), where tropical soils would be mineral alone, bereft of visible humus. But bringing back these heavy samples was a nightmare, not least because it had to be done without delay, before microbial activity influenced decay rates and the release of nitrogen and other salts into solution. Everything—everything—had to be kept waterproof, in camp and in transit, and this required commissioning special durable waterproof backpacks.
This work became my ecological specialty. We ended up with 105 plots, each 1.5 acres (increased from the Brunei experience), at thirteen sites in eleven localities. The work at each site, following establishment, which I personally undertook, was carried out by my team of climbers, some of whom had rejoined me when they heard of my return, and was led by an experienced Sarawak Malay forester. Among these foresters, I most remember Ilias bin Pa’ie, keeper of the Sarawak herbarium, who was a close friend and mentor, ever cautious and gentle, who tragically died from a heart attack when overseeing the 1975 recensusing at Lambir, a hilly site in the northeastern corner of the state. While this was some years after my departure, the loss was profound.
~
Otherwise, in those initial years, camp life during surveys became routine, with little of excitement to report. One exception was the accident our team experienced on the upper basalt slopes of Bukit Mersing, a mountain in central Sarawak where thirty plots were eventually installed, four of which became permanent for periodic recensusing. The Bornean climate is almost windless, except 00for the squalls that foreshadow the frequent afternoon thunderstorms. Occasionally, these take the form of violent cold-air downdrafts, flattening the forest in a patch of fifty acres or more. Our camp found itself in one when a giant emergent tree toppled nearby. The team hid beneath its cylindrical trunk, while the camp itself, including the tent frame and its tarpaulin, were trashed. But nothing was lost, and the work could continue.






And there were the occasional culinary surprises. We discovered, to our amazement, that civets at some camps would bite into the cans of tinned mackerel and suck out their contents. How did they know what was within? We deduced that a smell, or dry juice, had been left outside during the canning process. And, then again, there was the jaoung: Bornean forests have few canopy palm species, and most are scattered or local. This one is a Pholidocarpus, which locally formed small groves in damp valleys. It would be felled on discovery and cut open to reveal the massive starchy pith with the portly grubs of a large beetle nestled inside. The trick was to pick these up by the head and bite off the wriggling body, which resembled a greasy polythene tube full of shortening. Then, you had to swallow fast while the three pairs of scratchy legs tickled past your uvula. For me, once was enough!
~
We ultimately conducted four recensuses on five-year intervals, but the work still continues as I write. Different rainforest species achieve trunk-diameter growth rates from ten millimeters to less than two millimeters per annum, which is similar to a stand of regenerating red oaks (Quercus rubra) in a Massachusetts forest. The majority of individuals in any mature MDF, however, are losing to competitors and in slow decline. It required twenty years to gain sufficient data to start comparing the dynamic performance of forests on contrasting rock and soils, and then infer potential lengths of felling cycles and means of sustainable management for timber. By that time, I had spent twelve years on the biology faculty of the University of Aberdeen, where tropical forest research of another kind was occurring, and the work continued after I joined the Arnold Arboretum in 1978. It would take longer still before Harvard graduate student Matthew Potts was to use our data in his doctoral dissertation and devise the most informative methods to analyze patterns and correlations.

But even the first results were impressive. Our 105 plots covered 157 acres and included just under two hundred dipterocarp species exceeding one-foot girth. This amounted to four-fifths of all dipterocarps known from Sarawak and Brunei, even though our plots only included MDFs and therefore excluded the characteristic species of the peat swamp forest and other specialized communities. We even captured 70 percent of approximately thirty-five hundred known tree species in all families. This is tribute to our careful selection of sites representative of the full range of predicted species-specific habitats.
These initial results were amplified by Matthew, who devised an elegant method of visually depicting the similarities (or dissimilarities) between the plots. The model, known as a dendrogram, resembles a family tree, with the twigs and branches grouping plots according to their similarities. His analysis revealed several patterns of importance. First, the plots were grouped into two major branches, confirming the original observations and ordinations from Brunei: Fifty-seven Sarawak plots, primarily at eight sites, occurred on sandy soils, characterized by higher acidity, lower nutrient levels, and a distinct surface layer of slow-decomposing raw organic matter. The other forty-eight plots, mainly at the other five sites, occupied the more widespread fertile loams, with a higher capacity to retain water thanks to their open lattice of clay molecules. These major groups were consistent, regardless of the underlying bedrock (sedimentary or igneous).
Based on Matthew’s dendrogram, it could be argued that the reason why two-thirds of the species differed in our original Bruneian plots was simply because the distance between the two sites—sixty miles—is sufficient for the random turnover that might be expected over time from their restricted seed dispersal distances. But the plot sites from Sarawak correlated with soil properties, irrespective of their location, indicating that the selective influence of the physical habitat dominates the random effect of seed dispersal over time in structuring the tree communities. This is particularly evident for isolated plots, where the soil type differs from the surrounding forest, supporting a floristic island of tree species that differ from the surrounding habitat sea. Immediately, I realized that the limited dispersal of pollen and seeds results in tropical tree species evolving more-restricted, habitat-defined spaces through competitive speciation. These floristic islands form the ecological equivalent of terrestrial archipelagoes. Other organisms such as symbiotic fungi or insects whose larvae specifically depend on those trees will be similarly confined. The implications for conservation planning are obvious.
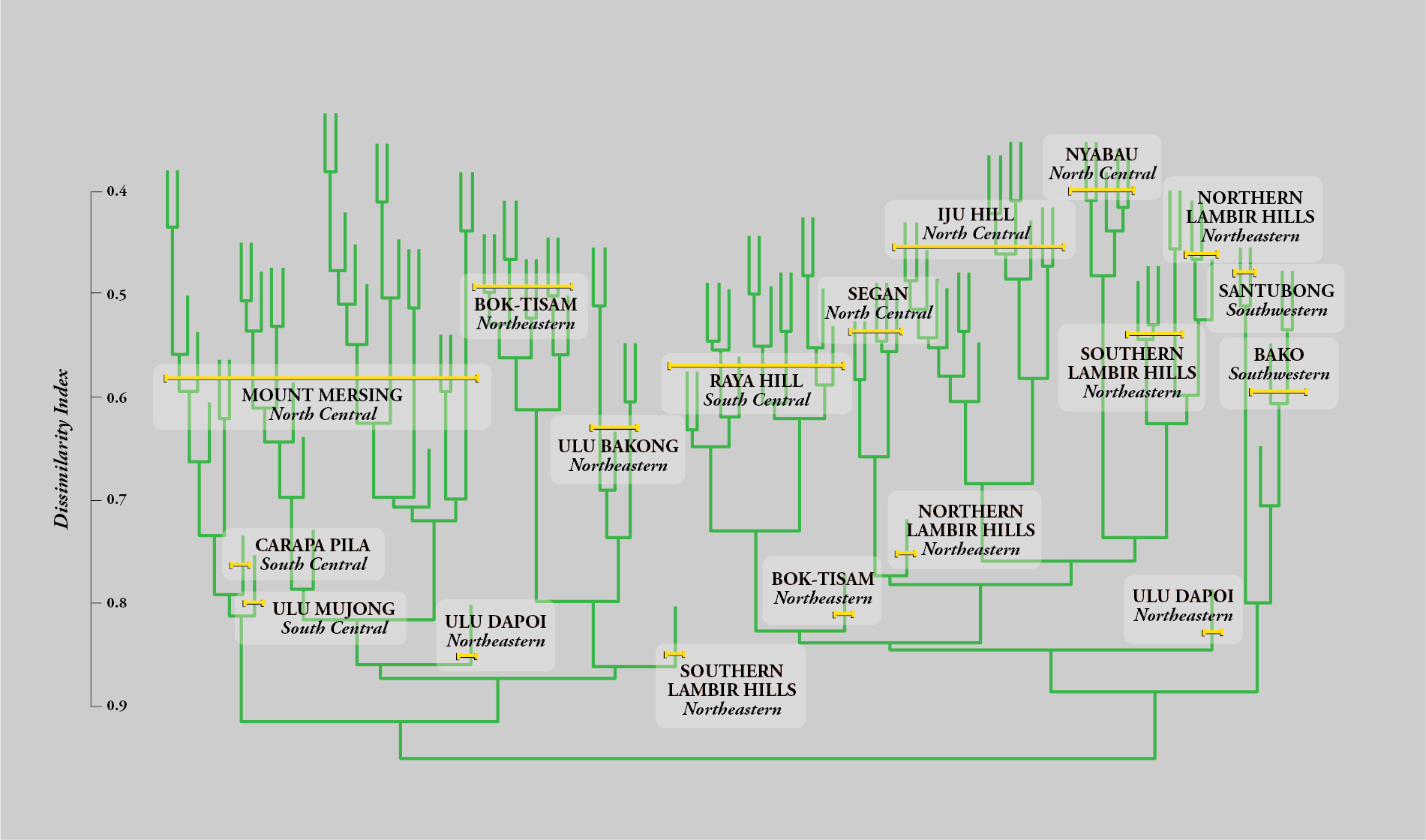
But although the plots within a particular site occupied distinct terminal branches of the dendrogram (no plots were identical), plots from a particular site generally remained within a single subsection of the dendrogram. This was even true for neighboring sites like the Bok-Tisam Forest Reserve and Ulu Bakong, which are located about twenty miles apart in northeastern Sarawak. Even though these sites share geology and soil characteristics (both primarily occur on yellow-brown loam), their forest compositions could be distinguished from one another. This provided strong evidence that, whereas soil properties dominate at broad landscape scales, the effects of dispersal are dominant over shorter distances, provided the soils are uniform. Within a single site, we also found that the floristic composition of the plots correlated with topography, as well as nutrient concentrations in the soil. Topography and nutrient concentrations were themselves interrelated, but we were eventually able, by clever analysis, to find that nutrient influences are generally stronger.
~
This research ultimately provided the basis for sustainable management of Bornean MDFs for timber production. Foresters in Peninsular Malaysia have never conducted plot surveys aimed at defining floristically distinct forest types, but they had, over many decades, developed a means of sustainably harvesting timber from MDFs by simulating natural succession: The forest is clearcut in patches hardly larger than large windthrows, after first checking to confirm that there was sufficient natural regeneration that would survive the logging process. Felling cycles of fifty to seventy years were envisaged. Our recensuses confirmed that Bornean forests on loam soils could be managed using the same method.

But foresters from Peninsular Malaysia are still finding it difficult to develop sustainable management systems for their high-hill and coastal dipterocarp forests, which floristically resemble the Bornean MDFs of low-nutrient sandy soils. Our findings have suggested a more sophisticated management system is necessary for these forests. Our forest structure measurements and recorded growth rates on these indicated that species attaining timber diameters were slow growing and in lower numbers, although there was an adequate cohort of young trees, rather than seedlings, for successive crops. Sustainable management would, therefore, require foresters return to the same stand at shorter intervals to selectively harvest individual trees, given that it might take a century for a seedling to mature to timber size. For this method, a more experienced workforce would be essential.
Our research also supported a new protocol for identifying the locations—to be tested by field censuses—of areas with potentially outstanding species diversity or concentration of endemics, meriting strict conservation. In this way, previously unexplored areas of conservation importance can be identified due to their surface geology. Up to then, conservationists were unable to make such extrapolations. Three national parks in Sarawak had already been legislated in the 1960s. I proposed five more, aimed at comprehensive representation of the flora. These were successfully passed into legislation by the new government of an independent Sarawakian state within Malaysia, in the 1970s, long after our departure from those raucous longhouse days and our entrance into the aethereal realm of academia.
Additional reading:
Ashton, P. S. 1964. Ecological studies in the mixed dipterocarp forests of Brunei State (Oxford forestry memoirs, no. 25). Oxford: Clarendon Press.
Ashton, P. S. 1982. Dipterocarpaceae. In C. G. G. J. van Steenis (Ed.), Flora Malesiana (Series 1, vol. 9, pp. 237–552). The Hague: Matinus Nijhoff.
Ashton, P. S. and Hall, P. 1992. Comparisons of structure among mixed dipterocarp forests of north-western Borneo. Journal of Ecology, 80: 459–491.
Ashton, P. S. 2014. On the forests of tropical Asia: Lest the memory fade. Richmond, Surrey: The Royal Botanic Gardens, Kew, in association with the Arnold Arboretum of Harvard University.
Beccari, O. 1904. Wanderings in the great forests of Borneo. F. H. H. Guillemard (Ed.) and E. H. Giglioli (Trans.). London: Archibald Constable and Co. (Original work published 1902.)
MacArthur, R. H. and Wilson, E. O. 1967. The theory of island biogeography (Monographs in population biology). Princeton: Princeton University Press.
Potts, M. D., Ashton, P. S., Kaufman, L. S. and Plotkin, J. B. 2002. Habitat patterns in tropical rain forests: A comparison of 105 plots in northwest Borneo. Ecology, 83(10): 2782–2797.
Peter Ashton is Harvard University Bullard Professor Emeritus and was director of the Arnold Arboretum from 1978 to 1987. Among many career honors, his research on tropical forests was recognized with the prestigious Japan Prize in 2007. He and his wife, Mary, live in Chiswick, London.
The map in this article was created using Esri, HERE, Garmin, (c) OpenStreetMap contributors, Sources: Esri, USGS, NGA, NASA, CGIAR, N Robinson, NCEAS, NLS, OS, NMA, Geodatastyrelsen, Rijkswaterstaat, GSA, Geoland, FEMA, Intermap and the GIS user community.
Citation: Ashton, P. 2019. A Botanist in Borneo: Understanding Patterns in the Forested Landscape. Arnoldia, 77(1): 26–39.